Lekársky expert článku

Nové publikácie
hypofýza
Posledná kontrola: 04.07.2025

Všetok obsah iLive je lekársky kontrolovaný alebo kontrolovaný, aby sa zabezpečila čo najväčšia presnosť faktov.
Máme prísne smernice týkajúce sa získavania zdrojov a len odkaz na seriózne mediálne stránky, akademické výskumné inštitúcie a vždy, keď je to možné, na lekársky partnerské štúdie. Všimnite si, že čísla v zátvorkách ([1], [2] atď.) Sú odkazmi na kliknutia na tieto štúdie.
Ak máte pocit, že niektorý z našich obsahov je nepresný, neaktuálny alebo inak sporný, vyberte ho a stlačte kláves Ctrl + Enter.
Hypofýza (hypofýza, s.glandula pituitaria) sa nachádza v hypofyzeálnej jamke tureckého sella klinovitej kosti a je oddelená od lebečnej dutiny výbežkom tvrdej mozgovej membrány, ktorý tvorí bránicu sella. Otvorom v tejto bránici je hypofýza spojená s infundibulom hypotalamu diencephalonu. Priečny rozmer hypofýzy je 10-17 mm, predozadný 5-15 mm, vertikálny 5-10 mm. Hmotnosť hypofýzy u mužov je približne 0,5 g, u žien 0,6 g. Hypofýza je zvonku pokrytá kapsulou.
V súlade s vývojom hypofýzy z dvoch rôznych zárodkov sa v orgáne rozlišujú dva laloky - predný a zadný. Adenohypofýza alebo predný lalok (adenohypofýza, s.lobus anterior) je väčší a tvorí 70 – 80 % celkovej hmotnosti hypofýzy. Je hustejší ako zadný lalok. V prednom laloku sa rozlišuje distálna časť (pars distalis), ktorá zaberá prednú časť jamky hypofýzy, stredná časť (pars intermedia), ktorá sa nachádza na hranici so zadným lalokom, a hľuznatá časť (pars tuberalis), ktorá smeruje nahor a spája sa s infundibulom hypotalamu. Vďaka množstvu krvných ciev má predný lalok svetložltú farbu s červenkastým odtieňom. Parenchým prednej hypofýzy je reprezentovaný niekoľkými typmi žľazových buniek, medzi ktorých vláknami sa nachádzajú sínusoidné krvné kapiláry. Polovica (50 %) buniek adenohypofýzy sú chromofilné adenocyty, ktoré majú vo svojej cytoplazme jemnozrnné granule, ktoré sa dobre farbia soľami chrómu. Ide o acidofilné adenocyty (40 % všetkých buniek adenohypofýzy) a bazofilné adenocyty (10 %). Medzi bazofilné adenocyty patria gonadotropné, kortikotropné a tyreotropné endokrinocyty. Chromofóbne adenocyty sú malé, majú veľké jadro a malé množstvo cytoplazmy. Tieto bunky sa považujú za prekurzory chromofilných adenocytov. Zvyšných 50 % buniek adenohypofýzy sú chromofóbne adenocyty.
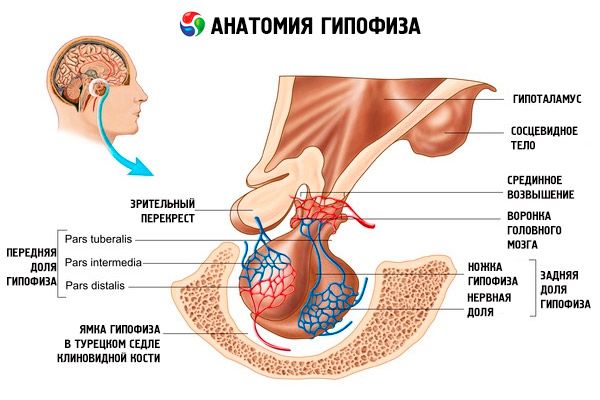
Neurohypofýza alebo zadný lalok (neurohypophysis, s.lobus posterior) pozostáva z nervového laloku (lobus nervosus), ktorý sa nachádza v zadnej časti jamy hypofýzy, a lievika (infundibulum), ktorý sa nachádza za hľuznatou časťou adenohypofýzy. Zadný lalok hypofýzy je tvorený neurogliálnymi bunkami (bunkami hypofýzy), nervovými vláknami, ktoré idú z neurosekrečných jadier hypotalamu do neurohypofýzy, a neurosekrečnými telieskami.
Hypofýza je prostredníctvom nervových vlákien (dráh) a krvných ciev funkčne prepojená s hypotalamom (diencephalon), ktorý reguluje činnosť hypofýzy. Hypofýza a hypotalamus spolu s ich neuroendokrinnými, cievnymi a nervovými spojeniami sa zvyčajne považujú za hypotalamo-hypofyzárny systém.
Hormóny prednej a zadnej hypofýzy ovplyvňujú mnohé funkcie tela, predovšetkým prostredníctvom iných žliaz s vnútornou sekréciou. V prednej hypofýze produkujú acidofilné adenocyty (alfa bunky) somatotropný hormón (rastový hormón), ktorý sa podieľa na regulácii rastových a vývojových procesov u mladých organizmov. Kortikotropné endokrinocyty vylučujú adrenokortikotropný hormón (ACTH), ktorý stimuluje vylučovanie steroidných hormónov nadobličkami. Tyreotropné endokrinocyty vylučujú tyreotropný hormón (TSH), ktorý ovplyvňuje vývoj štítnej žľazy a aktivuje produkciu jej hormónov. Gonadotropné hormóny: folikuly stimulujúci (FSH), luteinizačný (LH) a prolaktín - ovplyvňujú pohlavné dozrievanie tela, regulujú a stimulujú vývoj folikulov vo vaječníkoch, ovuláciu, rast mliečnych žliaz a produkciu mlieka u žien, proces spermatogenézy u mužov. Tieto hormóny produkujú bazofilné adenocyty (beta bunky). Vylučujú sa tu aj lipotropné faktory hypofýzy, ktoré ovplyvňujú mobilizáciu a využitie tukov v tele. V strednej časti predného laloku sa tvorí melanocyty stimulujúci hormón, ktorý riadi tvorbu pigmentov - melanínov - v tele.
Neurosekrečné bunky supraoptických a paraventrikulárnych jadier v hypotalame produkujú vazopresín a oxytocín. Tieto hormóny sú transportované do buniek zadnej hypofýzy pozdĺž axónov, ktoré tvoria hypotalamo-hypofyzárny trakt. Zo zadnej hypofýzy sa tieto látky dostávajú do krvi. Hormón vazopresín má vazokonstrikčný a antidiuretický účinok, pre ktorý sa nazýva aj antidiuretický hormón (ADH). Oxytocín má stimulačný účinok na kontraktilitu svalov maternice, zvyšuje sekréciu mlieka laktujúcou mliečnou žľazou, inhibuje vývoj a funkciu žltého telieska a ovplyvňuje zmeny v tonuse hladkých (nepriečne pruhovaných) svalov gastrointestinálneho traktu.
Vývoj hypofýzy
Predný lalok hypofýzy sa vyvíja z epitelu dorzálnej steny ústneho zálivu vo forme prstencovitého výrastku (Rathkeho vak). Tento ektodermálny výbežok rastie smerom k dnu budúcej tretej komory. Smerom k nemu, zo spodného povrchu druhého mozgového vačku (budúce dno tretej komory), vyrastá výbežok, z ktorého sa vyvíja sivý tuberkul infundibula a zadný lalok hypofýzy.
Cievy a nervy hypofýzy
Z vnútorných krčných tepien a ciev arteriálneho kruhu veľkého mozgu smerujú horná a dolná hypofyzárna tepna do hypofýzy. Horné hypofyzárne tepny idú do sivého jadra a infundibula hypotalamu, tu sa navzájom anastomózujú a tvoria kapiláry prenikajúce do mozgového tkaniva - primárna hemokapilárna sieť. Z dlhých a krátkych slučiek tejto siete sa tvoria portálne žily, ktoré smerujú do predného laloku hypofýzy. V parenchýme predného laloku hypofýzy sa tieto žily rozdeľujú na široké sínusoidné kapiláry a tvoria sekundárnu hemokapilárnu sieť. Zadný lalok hypofýzy je zásobovaný krvou hlavne dolnou hypofyzárnou tepnou. Medzi hornou a dolnou hypofyzárnou tepnou sú dlhé arteriálne anastomózy. Odtok venóznej krvi zo sekundárnej hemokapilárnej siete sa uskutočňuje systémom žíl, ktoré ústia do kavernóznych a interkavernóznych dutín dura mater mozgu.
Sympatické vlákna, ktoré vstupujú do orgánu spolu s tepnami, sa podieľajú na inervácii hypofýzy. Postgangliové sympatické nervové vlákna odchádzajú z plexu vnútornej krčnej tepny. Okrem toho sa v zadnom laloku hypofýzy nachádzajú početné zakončenia procesov neurosekrečných buniek nachádzajúcich sa v jadrách hypotalamu.
Vekom súvisiace znaky hypofýzy
Priemerná hmotnosť hypofýzy u novorodencov dosahuje 0,12 g. Hmotnosť orgánu sa do 10 rokov zdvojnásobí a do 15 rokov strojnásobí. Do 20 rokov hmotnosť hypofýzy dosahuje svoje maximum (530 – 560 mg) a v nasledujúcich vekových obdobiach zostáva takmer nezmenená. Po 60 rokoch sa pozoruje mierny pokles hmotnosti tejto endokrinnej žľazy.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Hormóny hypofýzy
Jednota nervovej a hormonálnej regulácie v tele je zabezpečená úzkym anatomickým a funkčným prepojením hypofýzy a hypotalamu. Tento komplex určuje stav a fungovanie celého endokrinného systému.
Hlavnou endokrinnou žľazou, ktorá produkuje množstvo peptidových hormónov priamo regulujúcich funkciu periférnych žliaz, je hypofýza. Je to červenosivý útvar v tvare fazule pokrytý vláknitou kapsulou s hmotnosťou 0,5-0,6 g. Jeho veľkosť sa mierne líši v závislosti od pohlavia a veku človeka. Všeobecne akceptované rozdelenie hypofýzy na dva laloky, ktoré sa líšia vývojom, štruktúrou a funkciou, zostáva: predný distálny - adenohypofýza a zadný - neurohypofýza. Prvý tvorí približne 70 % celkovej hmotnosti žľazy a konvenčne sa delí na distálnu, infundibulárnu a strednú časť, druhý - na zadnú časť alebo lalok a stopku hypofýzy. Žľaza sa nachádza v jamke hypofýzy turckého sedla klinovej kosti a je spojená s mozgom cez stopku. Horná časť predného laloku je pokrytá optickým chiazmom a optickými cestami. Krvné zásobenie hypofýzy je veľmi hojné a zabezpečujú ho vetvy vnútornej krčnej tepny (horná a dolná hypofyzárna tepna), ako aj vetvy arteriálneho kruhu mozgu. Horné hypofyzárne tepny sa podieľajú na krvnom zásobení adenohypofýzy a dolné - neurohypofýzy, pričom sa dotýkajú neurosekrečných zakončení axónov veľkých bunkových jadier hypotalamu. Prvé z nich vstupujú do strednej eminencie hypotalamu, kde sa rozptyľujú do kapilárnej siete (primárny kapilárny plexus). Tieto kapiláry (s ktorými sa dotýkajú zakončenia axónov malých neurosekrečných buniek mediobazálneho hypotalamu) sa zhromažďujú do portálnych žíl zostupujúcich pozdĺž stopky hypofýzy do parenchýmu adenohypofýzy, kde sa opäť delia do siete sínusových kapilár (sekundárny kapilárny plexus). Krv teda po predchádzajúcom prechode cez strednú vyvýšeninu hypotalamu, kde je obohatená o hypotalamické adenohypofyzotropné hormóny (uvoľňujúce hormóny), vstupuje do adenohypofýzy.
Odtok krvi nasýtenej adenohypofyzárnymi hormónmi z mnohých kapilár sekundárneho plexu sa uskutočňuje cez systém žíl, ktoré zase prúdia do venóznych dutín dura mater a potom do celkového krvného obehu. Portálny systém hypofýzy so zostupným smerom prietoku krvi z hypotalamu je teda morfofunkčnou súčasťou komplexného mechanizmu neurohumorálnej kontroly tropických funkcií adenohypofýzy.
Hypofýza je inervovaná sympatickými vláknami, ktoré sledujú hypofyzárne artérie. Vychádzajú z postgangliových vlákien, ktoré prechádzajú cez vnútorný karotický plexus a sú spojené s hornými cervikálnymi gangliami. Adenohypofýza nie je priamo inervovaná hypotalamom. Zadný lalok prijíma nervové vlákna z neurosekrečných jadier hypotalamu.
Adenohypofýza je svojou histologickou architektúrou veľmi zložitý útvar. Má dva typy žľazových buniek - chromofóbne a chromofilné. Tie sa zase delia na acidofilné a bazofilné (podrobný histologický popis hypofýzy je uvedený v príslušnej časti manuálu). Treba však poznamenať, že hormóny produkované žľazovými bunkami, ktoré tvoria parenchým adenohypofýzy, sa v dôsledku ich rozmanitosti do istej miery líšia svojou chemickou povahou a jemná štruktúra sekretujúcich buniek musí zodpovedať biosyntetickým vlastnostiam každej z nich. Niekedy však v adenohypofýze možno pozorovať prechodné formy žľazových buniek, ktoré sú schopné produkovať niekoľko hormónov. Existujú dôkazy o tom, že typ žľazových buniek adenohypofýzy nie je vždy určený geneticky.
Pod bránicou tureckého sedla sa nachádza lievikovitá časť predného laloku. Objíma stopku hypofýzy a dotýka sa sivého tuberkulu. Táto časť adenohypofýzy sa vyznačuje prítomnosťou epitelových buniek a bohatým prekrvením. Je tiež hormonálne aktívna.
Stredná (prostredná) časť hypofýzy pozostáva z niekoľkých vrstiev veľkých sekrečne aktívnych bazofilných buniek.
Hypofýza vykonáva rôzne funkcie prostredníctvom svojich hormónov. Jej predný lalok produkuje adrenokortikotropný (ACTH), tyreostimulačný (TSH), folikulystimulačný (FSH), luteinizačný (LH), lipotropný hormón, ako aj rastový hormón - somatotropný (STO) a prolaktín. V strednom laloku sa syntetizuje melanocyty stimulujúci hormón (MSH) a v zadnom laloku sa hromadí vazopresín a oxytocín.
ACTH
Hormóny hypofýzy sú skupinou proteínových a peptidových hormónov a glykoproteínov. Z hormónov prednej hypofýzy je ACTH najlepšie preskúmaný. Produkujú ho bazofilné bunky. Jeho hlavnou fyziologickou funkciou je stimulácia biosyntézy a sekrécie steroidných hormónov kôrou nadobličiek. ACTH tiež vykazuje melanocyty stimulujúcu a lipotropnú aktivitu. V roku 1953 bol izolovaný v čistej forme. Neskôr bola stanovená jeho chemická štruktúra, ktorá u ľudí a mnohých cicavcov pozostáva z 39 aminokyselinových zvyškov. ACTH nemá druhovú špecifickosť. V súčasnosti sa uskutočnila chemická syntéza samotného hormónu aj rôznych fragmentov jeho molekuly, ktoré sú aktívnejšie ako prirodzené hormóny. Štruktúra hormónu má dve časti peptidového reťazca, z ktorých jedna zabezpečuje detekciu a väzbu ACTH na receptor a druhá poskytuje biologický účinok. Zrejme sa viaže na receptor ACTH interakciou elektrických nábojov hormónu a receptora. Úlohu biologického efektora ACTH plní fragment molekuly 4-10 (Met-Glu-His-Phen-Arg-Tri-Tri).
Melanocyty stimulujúca aktivita ACTH je spôsobená prítomnosťou N-terminálnej oblasti v molekule, ktorá pozostáva z 13 aminokyselinových zvyškov a opakuje štruktúru alfa-melanocyty stimulujúceho hormónu. Táto oblasť tiež obsahuje heptapeptid prítomný v iných hypofyzárnych hormónoch a má určité adrenokortikotropné, melanocyty stimulujúce a lipotropné účinky.
Za kľúčový moment v účinku ACTH sa považuje aktivácia enzýmu proteínkinázy v cytoplazme za účasti cAMP. Fosforylovaná proteínkináza aktivuje enzým esterázu, ktorá premieňa estery cholesterolu na voľnú látku v tukových kvapôčkach. Bielkovina syntetizovaná v cytoplazme v dôsledku fosforylácie ribozómov stimuluje väzbu voľného cholesterolu na cytochróm P-450 a jeho prenos z lipidových kvapôčok do mitochondrií, kde sú prítomné všetky enzýmy, ktoré zabezpečujú premenu cholesterolu na kortikosteroidy.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Hormón stimulujúci štítnu žľazu
TSH - tyreotropín - hlavný regulátor vývoja a fungovania štítnej žľazy, procesov syntézy a sekrécie hormónov štítnej žľazy. Tento komplexný proteín - glykoproteín - pozostáva z alfa a beta podjednotiek. Štruktúra prvej podjednotky sa zhoduje s alfa podjednotkou luteinizačného hormónu. Navyše sa do značnej miery zhoduje u rôznych druhov zvierat. Poradie aminokyselinových zvyškov v beta podjednotke ľudského TSH bolo dešifrované a pozostáva zo 119 aminokyselinových zvyškov. Možno poznamenať, že beta podjednotky ľudského a hovädzieho TSH sú si v mnohých ohľadoch podobné. Biologické vlastnosti a povaha biologickej aktivity glykoproteínových hormónov sú určené beta podjednotkou. Zabezpečuje tiež interakciu hormónu s receptormi v rôznych cieľových orgánoch. Beta podjednotka však u väčšiny zvierat vykazuje špecifickú aktivitu až po spojení s alfa podjednotkou, ktorá pôsobí ako druh aktivátora hormónu. Tá s rovnakou pravdepodobnosťou indukuje luteinizačnú, folikuly stimulujúcu a tyreotropnú aktivitu určenú vlastnosťami beta podjednotky. Zistená podobnosť nám umožňuje dospieť k záveru, že tieto hormóny vznikli v procese evolúcie z jedného spoločného predchodcu, pričom beta podjednotka určuje aj imunologické vlastnosti hormónov. Existuje predpoklad, že alfa podjednotka chráni beta podjednotku pred pôsobením proteolytických enzýmov a tiež uľahčuje jej transport z hypofýzy do periférnych cieľových orgánov.
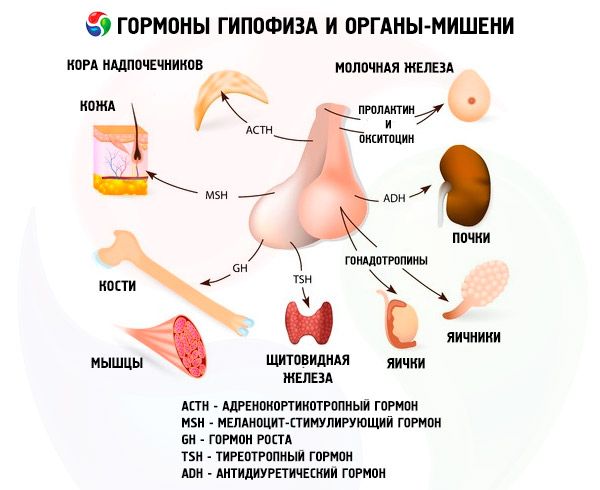
Gonadotropné hormóny
Gonadotropíny sú v tele prítomné vo forme LH a FSH. Funkčný účel týchto hormónov sa vo všeobecnosti obmedzuje na zabezpečenie reprodukčných procesov u jedincov oboch pohlaví. Rovnako ako TSH sú to komplexné proteíny - glykoproteíny. FSH indukuje dozrievanie folikulov vo vaječníkoch žien a stimuluje spermatogenézu u mužov. LH spôsobuje u žien prasknutie folikulov s tvorbou žltého telieska a stimuluje sekréciu estrogénov a progesterónu. U mužov ten istý hormón urýchľuje vývoj intersticiálneho tkaniva a sekréciu androgénov. Účinky gonadotropínov sú na sebe závislé a prebiehajú synchrónne.
Dynamika sekrécie gonadotropínov u žien sa mení počas menštruačného cyklu a bola dostatočne podrobne študovaná. V preovulačnej (folikulárnej) fáze cyklu je obsah LH na pomerne nízkej úrovni, zatiaľ čo FSH je zvýšený. S dozrievaním folikulu sa zvyšuje sekrécia estradiolu, čo podporuje zvýšenú produkciu gonadotropínov hypofýzou a výskyt cyklov LH aj FSH, t. j. pohlavné steroidy stimulujú sekréciu gonadotropínov.
V súčasnosti je štruktúra LH určená. Podobne ako TSH pozostáva z 2 podjednotiek: a a b. Štruktúra alfa podjednotky LH u rôznych živočíšnych druhov sa do značnej miery zhoduje, zodpovedá štruktúre alfa podjednotky TSH.
Štruktúra beta podjednotky LH sa výrazne líši od štruktúry beta podjednotky TSH, hoci má štyri identické úseky peptidového reťazca pozostávajúce zo 4-5 aminokyselinových zvyškov. V TSH sú lokalizované v pozíciách 27-31, 51-54, 65-68 a 78-83. Keďže beta podjednotka LH a TSH určuje špecifickú biologickú aktivitu hormónov, možno predpokladať, že homológne úseky v štruktúre LH a TSH by mali zabezpečovať spojenie beta podjednotiek s alfa podjednotkou a úseky, ktoré sa líšia štruktúrou, by mali byť zodpovedné za špecifickosť biologickej aktivity hormónov.
Natívny LH je veľmi stabilný voči pôsobeniu proteolytických enzýmov, ale beta podjednotka sa rýchlo štiepi chymotrypsínom a podjednotka a sa enzýmom ťažko hydrolyzuje, t. j. hrá ochrannú úlohu a bráni chymotrypsínu v prístupe k peptidovým väzbám.
Pokiaľ ide o chemickú štruktúru FSH, výskumníci zatiaľ nezískali definitívne výsledky. Podobne ako LH, aj FSH sa skladá z dvoch podjednotiek, ale beta podjednotka FSH sa líši od beta podjednotky LH.
Prolaktín
Ďalší hormón, prolaktín (laktogénny hormón), hrá aktívnu úlohu v reprodukčných procesoch. Hlavné fyziologické vlastnosti prolaktínu u cicavcov sa prejavujú vo forme stimulácie vývoja mliečnych žliaz a laktácie, rastu mazových žliaz a vnútorných orgánov. Podporuje prejav účinku steroidov na sekundárne pohlavné znaky u samcov, stimuluje sekrečnú aktivitu žltého telieska u myší a potkanov a podieľa sa na regulácii metabolizmu tukov. V posledných rokoch sa prolaktínu venuje veľká pozornosť ako regulátoru materského správania; takáto polyfunkčnosť sa vysvetľuje jeho evolučným vývojom. Je to jeden z najstarších hormónov hypofýzy a nachádza sa dokonca aj u obojživelníkov. V súčasnosti je štruktúra prolaktínu u niektorých druhov cicavcov úplne dešifrovaná. Vedci však až donedávna vyjadrovali pochybnosti o existencii takéhoto hormónu u ľudí. Mnohí sa domnievali, že jeho funkciu vykonáva rastový hormón. Teraz sa získali presvedčivé dôkazy o prítomnosti prolaktínu u ľudí a jeho štruktúra bola čiastočne dešifrovaná. Receptory prolaktínu sa aktívne viažu na rastový hormón a placentárny laktogén, čo naznačuje jeden mechanizmus účinku týchto troch hormónov.
Somatotropín
Rastový hormón, somatotropín, má ešte širšie spektrum účinku ako prolaktín. Rovnako ako prolaktín, aj on je produkovaný acidofilnými bunkami adenohypofýzy. STH stimuluje rast kostry, aktivuje biosyntézu bielkovín, má účinok mobilizácie tukov a podporuje zväčšenie telesnej veľkosti. Okrem toho koordinuje metabolické procesy.
Účasť hormónu v druhom prípade potvrdzuje skutočnosť prudkého zvýšenia jeho sekrécie hypofýzou, napríklad pri poklese hladiny cukru v krvi.
Chemická štruktúra tohto ľudského hormónu je teraz plne stanovená – 191 aminokyselinových zvyškov. Jeho primárna štruktúra je podobná štruktúre choriového somatomamotropínu alebo placentárneho laktogénu. Tieto údaje naznačujú významnú evolučnú blízkosť týchto dvoch hormónov, hoci vykazujú rozdiely v biologickej aktivite.
Je potrebné zdôrazniť vysokú druhovú špecifickosť daného hormónu - napríklad STH živočíšneho pôvodu je u ľudí neaktívny. Vysvetľuje sa to tak reakciou medzi receptormi STH ľudí a zvierat, ako aj samotnou štruktúrou hormónu. V súčasnosti prebiehajú štúdie zamerané na identifikáciu aktívnych centier v komplexnej štruktúre STH, ktoré vykazujú biologickú aktivitu. Študujú sa jednotlivé fragmenty molekuly, ktoré vykazujú iné vlastnosti. Napríklad po hydrolýze ľudského STH pepsínom bol izolovaný peptid pozostávajúci zo 14 aminokyselinových zvyškov a zodpovedajúci časti molekuly 31-44. Nemal rastový účinok, ale výrazne prekonal natívny hormón v lipotropnej aktivite. Ľudský rastový hormón, na rozdiel od podobného hormónu zvierat, má významnú laktogénnu aktivitu.
Adenohypofýza syntetizuje mnoho peptidových a proteínových látok, ktoré majú účinok mobilizácie tukov, a lipotropný účinok majú hypofyzárne tropné hormóny - ACTH, STH, TSH a ďalšie. V posledných rokoch sa osobitne zdôrazňujú beta- a γ-lipotropné hormóny (LPG). Biologické vlastnosti beta-LPG boli študované najpodrobnejšie; okrem lipotropnej aktivity má aj melanocyty stimulujúci, kortikotropín stimulujúci a hypokalcemický účinok a tiež vyvoláva inzulínu podobný účinok.
V súčasnosti je dešifrovaná primárna štruktúra ovčieho LPG (90 aminokyselinových zvyškov), lipotropných hormónov ošípaných a hovädzieho dobytka. Tento hormón má druhovú špecifickosť, hoci štruktúra centrálnej oblasti beta-LPG je u rôznych druhov rovnaká. To určuje biologické vlastnosti hormónu. Jeden z fragmentov tejto oblasti sa nachádza v štruktúre alfa-MSH, beta-MSH, ACTH a beta-LPG. Predpokladá sa, že tieto hormóny vznikli z rovnakého prekurzora v procese evolúcie. γ-LPG má slabšiu lipotropnú aktivitu ako beta-LPG.
Melanocyty stimulujúci hormón
Tento hormón, syntetizovaný v strednom laloku hypofýzy, stimuluje biosyntézu kožného pigmentu melanínu v jeho biologickej funkcii, podporuje zväčšenie veľkosti a počtu pigmentových buniek melanocytov v koži obojživelníkov. Tieto vlastnosti MSH sa používajú pri biologickom testovaní hormónu. Existujú dva typy hormónu: alfa- a beta-MSH. Ukázalo sa, že alfa-MSH nemá druhovú špecifickosť a má rovnakú chemickú štruktúru u všetkých cicavcov. Jeho molekula je peptidový reťazec pozostávajúci z 13 aminokyselinových zvyškov. Beta-MSH má naopak druhovú špecifickosť a jeho štruktúra sa u rôznych zvierat líši. U väčšiny cicavcov pozostáva molekula beta-MSH z 18 aminokyselinových zvyškov a iba u ľudí je od aminokyselinového konca predĺžená o štyri aminokyselinové zvyšky. Treba poznamenať, že alfa-MSH má určitú adrenokortikotropnú aktivitu a jeho vplyv na správanie zvierat a ľudí bol teraz dokázaný.
Oxytocín a vazopresín
Vazopresín a oxytocín, ktoré sa syntetizujú v hypotalame, sa hromadia v zadnom laloku hypofýzy: vazopresín v neurónoch supraoptického jadra a oxytocín v paraventrikulačnom jadre. Potom sa prenášajú do hypofýzy. Treba zdôrazniť, že prekurzor hormónu vazopresínu sa syntetizuje najprv v hypotalame. Zároveň sa tam produkujú proteíny neurofyzín typu 1 a 2. Prvý sa viaže na oxytocín a druhý na vazopresín. Tieto komplexy migrujú vo forme neurosekrečných granúl v cytoplazme pozdĺž axónu a dosahujú zadný lalok hypofýzy, kde nervové vlákna končia v cievnej stene a obsah granúl vstupuje do krvi. Vazopresín a oxytocín sú prvé hormóny hypofýzy s úplne stanovenou aminokyselinovou sekvenciou. Podľa svojej chemickej štruktúry sú to nonapeptidy s jedným disulfidovým mostíkom.
Uvažované hormóny majú rôzne biologické účinky: stimulujú transport vody a solí cez membrány, majú vazopresorický účinok, zvyšujú kontrakcie hladkého svalstva maternice počas pôrodu a zvyšujú sekréciu mliečnych žliaz. Treba poznamenať, že vazopresín má vyššiu antidiuretickú aktivitu ako oxytocín, pričom oxytocín má silnejší účinok na maternicu a mliečnu žľazu. Hlavným regulátorom sekrécie vazopresínu je spotreba vody; v renálnych tubuloch sa viaže na receptory v cytoplazmatických membránach s následnou aktiváciou enzýmu adenylátcyklázy v nich. Za väzbu hormónu na receptor a za biologický účinok sú zodpovedné rôzne časti molekuly.
Hypofýza, spojená cez hypotalamus s celým nervovým systémom, spája endokrinný systém do funkčného celku a podieľa sa na zabezpečovaní stálosti vnútorného prostredia tela (homeostáza). V rámci endokrinného systému sa homeostatická regulácia vykonáva na princípe spätnej väzby medzi predným lalokom hypofýzy a „cieľovými“ žľazami (štítna žľaza, kôra nadobličiek, pohlavné žľazy). Nadbytok hormónu produkovaného „cieľovou“ žľazou inhibuje a jeho nedostatok stimuluje sekréciu a uvoľňovanie zodpovedajúceho tropného hormónu. Hypotalamus je súčasťou systému spätnej väzby. Práve v ňom sa nachádzajú receptorové zóny citlivé na hormóny „cieľových“ žliaz. Špecifickou väzbou na hormóny cirkulujúce v krvi a zmenou reakcie v závislosti od koncentrácie hormónov prenášajú hypotalamické receptory svoj účinok do zodpovedajúcich hypotalamických centier, ktoré koordinujú prácu adenohypofýzy a uvoľňujú hypotalamické adenohypofyziotropné hormóny. Hypotalamus by sa preto mal považovať za neuroendokrinný mozog.