
Vaječník
Posledná kontrola: 23.04.2024

Všetok obsah iLive je lekársky kontrolovaný alebo kontrolovaný, aby sa zabezpečila čo najväčšia presnosť faktov.
Máme prísne smernice týkajúce sa získavania zdrojov a len odkaz na seriózne mediálne stránky, akademické výskumné inštitúcie a vždy, keď je to možné, na lekársky partnerské štúdie. Všimnite si, že čísla v zátvorkách ([1], [2] atď.) Sú odkazmi na kliknutia na tieto štúdie.
Ak máte pocit, že niektorý z našich obsahov je nepresný, neaktuálny alebo inak sporný, vyberte ho a stlačte kláves Ctrl + Enter.
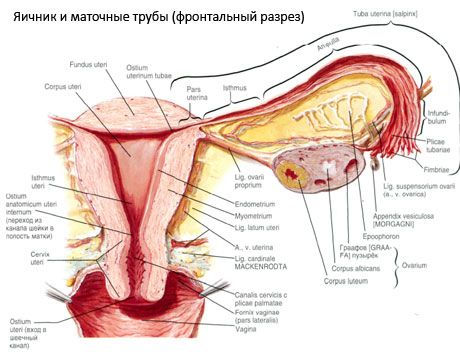
Vaječník (ovaria, grécky oophorón) - spárovaný orgán, ženské pohlavné žľazy, ktoré sa nachádzajú v dutine malej panvy za širokou väzbou maternice. Ovarie vyvíjajú a zraňujú samice pohlavných buniek (ovules), rovnako ako ženské pohlavné hormóny, ktoré vstupujú do krvi a lymfy. Vaječný vaječník má vajcovitý tvar, čiastočne sploštený v prednej a zadnej časti. Farba vaječníkov je ružová. Na povrchu vaječníkov ženy, ktorá rodí, sú viditeľné depresie a jazvy - stopy ovulácie a premeny žltých teliesok. Hmotnosť vaječníkov bol 8,5 g rozmery vaječníkov sú:. Dĺžka 2,5-5,5 cm, 1,5-3,0 cm šírka, hrúbka - až 2 cm sa ovariálny rozlišovať dva voľné povrchy: stredový povrch (facies medialis ) privrátenej k panvovej dutiny, čiastočne pokryté vajcovodu, a bočné povrch (facie lateralis), ktorá prilieha k panvovej bočnej steny, mierne prehĺbenie - ovariálne fossa. Táto fovea leží v rohu medzi peritoneálnymi vonkajšími iliacami v hornej časti a maternicovými a okluzívnymi artériami na dne. Za vaječníkom prechádza močovod z príslušnej strany zhora nadol.
Ovariálny povrch pohybujúce sa vo vypuklom zadarmo (zadná) hrany (margo Liber), na prednej strane - v mezenterické oblasti (Margo mesovaricus), pomocou krátkeho klipu násobne pobrušnice (okružie vaječníka) na zadnom liste širokej väzu maternice. V čele telesa je vrúbkovaný vybranie - vaječník brána (hilum ovárií), prostredníctvom ktorého vaječníkov patrí tepny a nervy ležia žily a lymfatických ciev. Ďalej izolované vaječníkov dva konce: zaoblenou horný rúrkový koniec (extremitas tubaria), naproti vajcovodu, a dolný koniec matky (extremitas utenna), v spojení s maternice vlastnou zväzok vaječníkov (líg ovárií proprium.). Tento balíček v okrúhle šnúry okolo 6 mm od matky je koniec k bočnej rohovej vaječníkov krčka, ktorá sa nachádza medzi týmito dvoma vrstvami širokého väzu. Tým, väzivového aparátu vaječníka sa tiež týka väz podveshivayaschaya vaječník (lig.suspensorium ovárií), ktorý je záhyb pobrušnice, siahajúce od hornej steny panvy na vaječník a v nádobách vaječníkov obsahujúce a vláknitých zväzkov vlákien. Vaječník pevné krátke mezentéria (mesovarium), čo predstavuje duplikatury pobrušnicu, rozprestierajúce sa od zadného cípe širokého väzu maternice na vaječníku mezenterické hrany. Samotné vaječníky nie sú kryté peritoneom. Najväčšia fimbria vaječníka je pripojená ku koncu vaječníka. Topografia vaječníkov závisí od polohy maternice, od jej veľkosti (počas tehotenstva). Ovarie sa týkajú veľmi pohyblivých orgánov panvovej dutiny.
 [
[Ovariálne cievy a nervy
Krvný obeh vaječníkov je spôsobený aa. A vv. Ovaricae a uterinae. (. Aa ovaricae Dextra et sinistra) Ako vaječníkov tepna sa rozkladajú od predného povrchu aorty tesne pod obličkové tepny, pravá najčastejšie pochádza z aorty a vľavo - renálnej artérie. Ísť dole a priečne cez prednú povrch psoas svalu, vaječníkov tepny pretína každú prednú močovodu (čo mu vetvičky), vonkajšie bedrové plavidiel a hraničnú čiaru prísť do panvovej dutiny, pretože sa nachádza tu v odkladný väzu vaječníku. Po mediálne, vaječníkov tepna prechádza medzi listami širokej väzu maternice za vajcovodov, čo je vetvy, a potom - v okružia vaječníku; vstúpi do brány vaječníkov.
Vetvy vaječníkovej tepny sú široko anastomózne s ovariálnymi ramenami maternice. Venózny výtok z vaječníkov sa uskutočňuje predovšetkým v ovariálnom venóznom plexu, ktorý sa nachádza v oblasti vaječníkov. Preto odtok krvi prechádza dvomi smermi: cez žily maternice a vaječníkov. Pravá ovariálna žila má ventily a prechádza do dolnej dutej žily. Ľavá vaječníková žila prúdi do ľavej obličkovej žily bez prítomnosti ventilov.
Lymfatický výtok z vaječníkov sa prejavuje cez lymfatické cievy, obzvlášť bohaté v oblasti orgánových brán, kde sa vylučuje sublingválny lymfatický plexus. Potom sa lymfa odkloní do para-aortálnych lymfatických uzlín pozdĺž vaječníkových lymfatických ciev.
Inervácia vaječníkov
Sympatika - je poskytovaná postgangliovými vláknami z celiakie (solárne), zvrchovaných a hypogastrických plexusov; parasympatiku - kvôli vnútorným sakrálnym nervom.
Štruktúra vaječníkov
Povrch vaječníkov je pokrytý jednovrstvovým germinálnym epitelom. Pod ním je hustá obálka spojivového tkaniva (tunica albuginea). Spojivové tkanivo vaječníka foriem stróma (stróma ovárií), bohaté na elastických vlákien. Podstata vaječníkov, jeho parenchým, je rozdelená na vonkajšie a vnútorné vrstvy. Vnútorná vrstva, ktorá leží v strede vaječníkov, bližšie k bráne, tzv dreň (medulla ovárií). V tejto vrstve, vo voľnej spojivového tkaniva sú početné krv a lymfatické cievy a nervy. Vonkajšia vrstva vaječníkov - kôra vaječníka je hustšia. To má mnoho spojivového tkaniva, v ktorých sú umiestnené dozrievania primárnych folikulov (folliculi ovarici primaria), sekundárne (bublín) folikulov (folliculi ovarici Secundaria, s.vesiculosi), A dobre zrelých folikulov Graafovho bubliny (folliculi ovarici maturis), a žltý a atretických teliesok.
V každom folikulu je samica reprodukčného vajíčka alebo oocytu (ovocytus). Priemer oocytov až 150 mikrónov, zaoblené, obsahuje jadro, veľké množstvo cytoplazmy, ktorý, okrem bunkových organel, existujú proteín-lipid inklúzie (žĺtok), glykogén potrebná pre zásobovanie vajec. Dodávka oocytov zvyčajne spotrebuje do 12 až 24 hodín po ovulácii. Ak nedôjde k oplodneniu, vajíčko zomrie.
Ľudský vaječník má dve krycie membrány. Vo vnútri je cytolema, čo je cytoplazmatická membrána oocytov. Mimo cytolemu sa nachádza vrstva takzvaných folikulárnych buniek, ktoré chránia vajíčko a vykonávajú funkciu tvorcu hormónov - uvoľňovanie estrogénu.
Fyziologická poloha maternice, vajíčok a vaječníkov je zabezpečená zavesením, upevnením a podporným zariadením. Kombinácia peritonea, väziva a panvovej celulózy. Závesné zariadenie je reprezentované spárovanými formáciami, obsahuje okrúhle a široké väzy maternice, vlastné väzy a zavesené vagíny. Široké väzivo maternice, vlastné a suspendujúce väzivo vaječníkov, udržujú maternicu v strednej polohe. Okrúhle väzy priťahujú dno maternice dopredu a poskytujú svoj fyziologický sklon.
Upevňovacie zariadenie zaisťuje polohu tiahnúcich sa v strede malej panvy a prakticky nemožno ich posúvať na strany, tam a späť. Pretože sa väzivo pohybuje od maternice v jeho spodnej časti, je možné nakloniť maternicu v rôznych smeroch. V upevňovacie jednotka obsahuje šnúry usporiadané do voľnej panvovej tkaniva a vystupujúce z dolnej maternice karty do boku, predné a zadné steny panvy: krížovodriekové magochnye Cardinal, maternice a cystická-Vezikovaginální lonovej väzu.
Okrem mesovaria sa rozlišujú nasledujúce ovariálne väzy:
- suspenzia vajcového väzu, predtým označovaná ako voronkotazovaya. Je záhyb peritonea s ňou prebiehajúcej krvi (a. Et v. Ovarica) a lymfatické cievy a nervy vaječníkov napnutý medzi bočnej stene panvové, bedrové fascie (v rozdelení spoločnej arteria iliaca na vonkajšie a vnútorné) a horné (trubky) koniec vaječníkov;
- ovariálne vlastné väzivo prechádza medzi vrstvami širokého maternicového väzba, bližšie k zadnému listu a spája dolný koniec vaječníkov s bočným okrajom maternice. Do maternice je vlastné väziva medzi vpichom maternice a kruhovým väzivom, ktoré sa navzájom spájajú. V hrúbke väzov sú rr. Ovarii, ktoré sú koncovými oblasťami maternice;
- uzáver apendikulárneho vaječníka Chlopňa sa rozprestiera okolo hrebeňa doplnku do pravého vaječníka alebo širokého väzu maternice vo forme záhybu peritonea. Väzivo je nestabilné a pozoruje sa u 1/2 až 1/3 žien.
Nosné zariadenie je reprezentované svalmi a fasciami panvového dna, rozdelené na spodnú, strednú a hornú (vnútornú) vrstvu.
Najsilnejšou je horná (vnútorná) svalová vrstva, ktorú predstavuje spárovaný sval, ktorý zvyšuje konečník. Skladá sa zo svalových zväzkov, ktoré sa vyvíjajú z kokyxu do panvových kostí v troch smeroch (pubik-kokcygeal, ilio-coccygeal a ischiococcygeal svaly). Táto vrstva svalov sa nazýva aj membrána panvy.
Stredná vrstva svalov je umiestnená medzi kosti symphysis, pubic a ischium. Stredná vrstva svalov - urogenitálna membrána - zaberá prednú polovicu panvového výstupu, prechádza cez uretru a vagínu. V prednej časti medzi jej listami sú svalové lúče, ktoré tvoria vonkajší zviera uretry, v zadnej časti sú svalové zväzky prechádzajúce v priečnom smere, hlboký priečny sval perinea.
Spodný (vonkajšie) vrstvy svalov panvového dna sa skladá z plochy, ktorá poloha je v tvare obrázku 8. Tieto zahŕňajú bulbospongiosus-kavernózna, zrastu ischio-kavernózna, vonkajší zvierač z konečníka, povrchné priečny perineálnej svaly.
Ontogénnosť vaječníkov
Proces rastu a folikulárnej atrézie začína s 20 týždňami tehotenstva a v čase dodania vo vaječníkoch dievčaťa zostáva až 2 milióny oocytov. V čase menarche sa ich počet znižuje na 300 tisíc.V priebehu celého obdobia reprodukčného života dosiahne zrelosť a ovuluje nie viac ako 500 folikulov. Počiatočný rast folikulov nezávisí od stimulácie FSH, je obmedzený a atrézia nastane rýchlo. Predpokladá sa, že namiesto steroidných hormónov sú miestne autokrinné / parakrínové peptidy hlavným regulátorom rastu a atrézie primárnych folikulov. Predpokladá sa, že proces rastu a atrézie folikulov nie je prerušený žiadnymi fyziologickými procesmi. Tento proces pokračuje v každom veku, vrátane intrauterinného obdobia a menopauzy, je prerušený tehotenstvom, ovuláciou a anovuláciou. Mechanizmus, ktorý spúšťa rast folikulov a ich počet v každom konkrétnom cykle, zatiaľ nie je jasný.
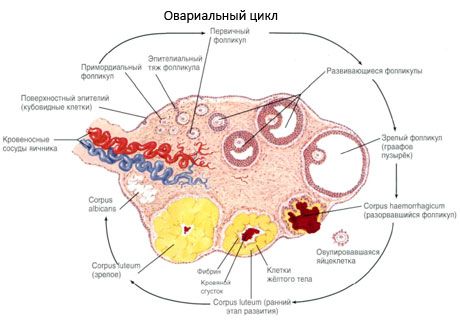
Vo svojom vývoji sa folikul podrobuje niekoľkým fázam vývoja. Primordiálne zárodočné bunky pochádzajú z endodermu žĺtkového vaku, allantois a migrujú do genitálnej oblasti embrya v 5. - 6. Týždni tehotenstva. Rýchly mitotické, ktorý sa rozprestiera od 6-8nedel 16-20nedel tehotenstva, fetálny vaječník tvorený 6-7 miliónov oocyty obklopených tenkou vrstvou granulosa buniek.
Predantrálny folikul - oocyt je obklopený membránou (Zona pellucida). Granulózne bunky obklopujúce oocyty začínajú proliferovať, ich rast závisí od gonadotropínov a koreluje s hladinou estrogénov. Granulózne bunky sú cieľom pre FSH. V štádiu predantrálneho folikulu môžu granulózne bunky syntetizovať tri triedy steroidov: prednostne indukujú aktivitu aromatázy, hlavného enzýmu, ktorý premieňa androgény na estradiol. Predpokladá sa, že estradiol je schopný zvýšiť počet svojich vlastných receptorov, čo poskytuje priamy mitogénny účinok na granulózne bunky nezávislé od FSH. Je považovaný za parakrinný faktor, ktorý zvyšuje účinky FSH vrátane aktivácie aromatizujúcich procesov.
FSH receptory sa objavujú na membránach granulózových buniek, hneď ako začne rast folikulu. Zníženie alebo zvýšenie hladiny FSH vedie k zmene počtu jeho receptorov. Toto pôsobenie FSH je modulované rastovými faktormi. FSH pôsobí prostredníctvom G-proteínu adenylát-cyklasy systému vo folikule steroidogenézy hoci najmä regulované FSH, proces zahŕňa mnoho faktorov: iónové kanály, receptorové tyrozínkinázy fosfolipázy systém sekundárnych poslov.
Úloha androgénov v skorom vývoji folikulov je zložitá. Bunky granulózy majú androgénne receptory. Nie sú substrátom iba pre FSH-indukovanej aromatizácia na estrogén, ale v nízkych koncentráciách môžu zlepšiť proces ochutenie. Keď je hladina androgénov zvyšuje preantral granulosa bunky prednostne vybrané cesty žiadne aromatizáciu do estrogénov a jednoduchší spôsob pre konverziu pomocou androgénov 5a-reduktázy v rozvojovom androgén, ktoré nemôžu byť prevedené na estrogén, a tým inhibovala aktivitu aromatázy. Tento proces tiež inhibuje tvorbu FSH a LH receptory, čím sa zastaví vývoj folikulu.
Proces aromatizácie, folikul s vysokou hladinou androgénov podlieha procesom atrézie. Rast a vývoj folikulu závisí od jeho schopnosti previesť androgény na estrogény.
V prítomnosti FSH je dominantnou látkou folikulárnej tekutiny estrogény. V neprítomnosti FSH - androgénov. LH je normálna vo folikulárnej tekutine až do polovice cyklu. Akonáhle sa zvýšená úroveň mitotickej aktivity granulosa bunky LH znižuje, a degeneratívne zmeny javí zvýšená hladina androgénu hladín steroidných folikul vo folikulárnej tekutine ako v plazme a odráža funkčnú aktivitu vaječník: granulosa a theca bunky. Ak je jediným cieľom pre FSH granulózne bunky, LH má mnoho cieľov - to sú bunky, stromálne a luteálne bunky a granulózne bunky. Schopnosť steroidogenézy má ako bunky granulózy, tak teka, ale aktivita aromatázy prevažuje v bunkách granulózy.
V reakcii na LH bunky teka produkujú androgény, ktoré potom prostredníctvom aromatizácie indukovanej FSH sú transformované bunkami granulózy na estrogény.
Ako folikulu theca bunky začnú exprimovat gény pre receptor LH P450 sec a 3bet-hydroxysteroiddehydrogenázou dehydrogenázy, rastového faktora podobného inzulínu (IGF-1) synergicky s LH pre zvýšenie expresie génu, ale nestimulujú steroidov.
Ovariálna steroidogenéza je vždy závislá od LH. Ako rastú folikul, súčasné bunky exprimujú enzým P450c17, ktorý tvorí androgén z cholesterolu. Bunky granulózy nemajú tento enzým a sú závislé od súčasných buniek pri produkcii estrogénov z androgénov. Na rozdiel od steroidogenézy - folikulogenéza závisí od FSH. Ako folikulu a zvyšujú hladinu estrogénu prichádza do akčného mechanizmu spätnej väzby - inhibujú produkciu FSH, čo vedie k zníženiu aktivity aromatázy folikulu a v konečnom dôsledku k folikuly atrézia cez apoptózu (programovanú bunkovú smrť).
Mechanizmus spätnej väzby estrogénov a FSH inhibuje vývoj folikulov, ktoré začali rásť, ale nie dominantný folikul. Dominantný folikul obsahuje viac FSH receptorov, ktoré podporujú proliferáciu granulózových buniek a aromatizáciu androgénov v estrogénoch. Okrem toho parakrinná a autokrinná dráha pôsobí ako dôležitý koordinátor vývoja antralového folikulu.
Neoddeliteľnou súčasťou autokrinného / parakrínového regulátora sú peptidy (inhibín, aktivín, follistatin), ktoré sú syntetizované bunkami granulózy v reakcii na pôsobenie FSH a vstupujú do folikulárnej tekutiny. Inhibín znižuje sekréciu FSH; aktivín stimuluje uvoľňovanie FSH z hypofýzy a zvyšuje účinok FSH vo vaječníku; Follistatin potláča aktivitu FSH, pravdepodobne v dôsledku väzby aktivínu. Po ovulácii a vývoji žltého tela je inhibín pod kontrolou LH.
Rast a diferenciácia ovariálnych buniek je ovplyvnená inzulínovými rastovými faktormi (IGE). IGF-1 pôsobí na bunky granulózy, čo spôsobuje zvýšenie cyklického adenozínmonofosfátu (cAMP), progesterónu, oxytocínu, proteoglykánu a inhibínu.
IGF-1 pôsobí na teka bunky, čo spôsobuje zvýšenie produkcie androgénov. Teka bunky zase produkujú faktor nekrózy nádorov (TNF) a epidermálny rastový faktor (EGF), ktoré sú tiež regulované FSH.
EGF stimuluje proliferáciu buniek granulózy. IGF-2 je hlavným faktorom rastu folikulárnej tekutiny, ale tiež detegoval IGF-1, TNF-a, TNF-3 a EGF.
Zdá sa, že porušenie parakrínovej a / alebo autokrinnej regulácie funkcie vaječníkov zohráva úlohu pri narušení ovulačných procesov a pri tvorbe polycystických vaječníkov.
S rastúcim antrálnym folikulom sa zvyšuje obsah estrogénov vo folikulárnej tekutine. Na vrchole ich nárastu na bunkách granulózy sa objavujú receptory LH, dochádza k luteinizácii granulóznych buniek a produkcia progesterónu sa zvyšuje. Preto počas predovulovacej periódy zvýšenie produkcie estrogénov spôsobuje výskyt LH receptorov, LH naopak spôsobuje luteinizáciu buniek granulózy a produkciu progesterónu. Zvýšenie progesterónu znižuje hladinu estrogénov, čo zjavne spôsobuje druhý vrchol FSH uprostred cyklu.
Predpokladá sa, že ovulácia nastáva 10-12 hodín po vrchole LH a 24-36 hodín po vrchole estradiolu. Predpokladá sa, že LH stimuluje redukciu oocytov, luteinizáciu buniek granulózy, syntézu progesterónu a prostaglandínu vo folikuloch.
Progesterón zvyšuje aktivitu proteolytických enzýmov spolu s prostaglandínom, ktorý sa podieľa na pretrhnutí steny folikulu. FSH vyvolané progesterónový pík, umožňuje výstup oocytu z folikulu prevedením plazminogén na proteolytický enzým - plazmín, poskytuje dostatočné množstvo LH receptora pre normálny vývoj luteálnej fázy.
Do 3 dní po ovulácii, granulosa bunky zvyšuje, sa objavujú charakteristické vakuoly naplnené pigment - luteín. Teka-luteálne bunky sa odlišujú od teki a stromy a stávajú sa súčasťou žltého tela. Veľmi rýchlo sa pod vplyvom angiogénnych faktorov je rozvoj kapilár, ktoré prestupujú žlté teliesko, ak zlepšeniu vaskularizácie zvýšená produkcia estrogénu a progesterónu. Steroidogenézy aktivita a dĺžka života žltého telieska sa určuje podľa úrovne LH. Žlté telo nie je homogénna bunková entita. Prídavok 2 typy luteálnej buniek obsahuje endotelové bunky, makrofágy, fibroblasty, a ďalšie. Veľká luteálnej bunky produkujú peptidy (relaxínu oxytocín) a sú aktívne vo steroidogenézy viac aktivitu aromatázy a veľká syntéza progesterónu ako malých buniek.
Pík progesterónu sa pozoroval 8. Deň po vrchole LG. Zistilo sa, že progesterón a estradiol v luteálnej fáze sú vylučované sporadicky v korelácii s pulzným výstupom LH. Pri tvorbe žltého tela kontrola produkcie inhibínu prechádza z FSH na LH. Ingibin sa zvyšuje so zvýšením estradiolu na vrchol LH a naďalej sa zvyšuje po vrchole LH, aj keď hladina estrogénov klesá. Aj keď sú inhibíny a estradiol sekretované bunkami granulózy, regulujú sa rôznymi spôsobmi. Pokles inhibítorov na konci luteálnej fázy prispieva k zvýšeniu FSH pre ďalší cyklus.
Žlté telo veľmi rýchlo - 9 až 11 dní po ovulácii klesá.
Mechanizmus degenerácie nie je jasný a nesúvisí s luteolitickou úlohou estrogénov alebo s mechanizmom spojeným s receptorom, ako je vidieť v endometriu. Existuje ďalšie vysvetlenie úlohy estrogénov produkovaných žltým telom. Je známe, že pre syntézu progesterónových receptorov v endometriu sú potrebné estrogény. Euterogénne fázové estrogény sú pravdepodobne potrebné pre zmeny súvisiace s progesterónom v endometriu po ovulácii. Nedostatočný vývoj progesterónových receptorov v dôsledku neadekvátneho obsahu estrogénu je pravdepodobne ďalším mechanizmom neplodnosti a skorého tehotenstva, inou formou menejcennosti luteálnej fázy. Predpokladá sa, že životnosť žltého tela je nastavená v čase ovulácie. A bude určite regresované, ak choriový gonadotropín nie je podporovaný v súvislosti s tehotenstvom. Regresia žltého tela vedie k zníženiu hladín estradiolu, progesterónu a inhibínu. Redukčný inhibítor odstraňuje svoj inhibičný účinok na FSH; redukcia estradiolu a progesterónu umožňuje veľmi rýchlo obnoviť sekréciu GnRH a odstrániť mechanizmus spätnej väzby z hypofýzy. Redukcia inhibínu a estradiolu spolu so zvýšením GnRH vedie k prevalencii FSH nad LH. Zvýšenie hladiny FSH vedie k rastu folikulov s následnou voľbou dominantného folikulu a začne nový cyklus v prípade, že tehotenstvo nenastane. Steroidné hormóny zohrávajú vedúcu úlohu v reprodukčnej biológii a všeobecne vo fyziológii. Určujú fenotyp osoby, ovplyvňujú kardiovaskulárny systém, metabolizmus kostí, pokožku, celkovú pohodu tela a zohrávajú kľúčovú úlohu v tehotenstve. Účinok steroidných hormónov odráža intracelulárne a genetické mechanizmy, ktoré sú potrebné na prenos extracelulárneho signálu do jadra bunky na vyvolanie fyziologickej odpovede.
Estrogény difundujú cez bunkovú membránu a viažu sa na receptory umiestnené v jadre bunky. Komplex receptor-steroid sa potom viaže na DNA. V cieľových bunkách tieto interakcie vedú k expresii génov, k syntéze proteínov, k špecifickej funkcii buniek a tkanív.