Lekársky expert článku

Nové publikácie
Syntéza, vylučovanie a metabolizmus katecholamínov
Posledná kontrola: 06.07.2025

Všetok obsah iLive je lekársky kontrolovaný alebo kontrolovaný, aby sa zabezpečila čo najväčšia presnosť faktov.
Máme prísne smernice týkajúce sa získavania zdrojov a len odkaz na seriózne mediálne stránky, akademické výskumné inštitúcie a vždy, keď je to možné, na lekársky partnerské štúdie. Všimnite si, že čísla v zátvorkách ([1], [2] atď.) Sú odkazmi na kliknutia na tieto štúdie.
Ak máte pocit, že niektorý z našich obsahov je nepresný, neaktuálny alebo inak sporný, vyberte ho a stlačte kláves Ctrl + Enter.
Dreň nadobličiek produkuje zlúčeniny so štruktúrou, ktorá sa veľmi podobá steroidom. Obsahujú 3,4-dihydroxyfenylové (katecholové) jadro a nazývajú sa katecholamíny. Patria sem adrenalín, noradrenalín a dopamín beta-hydroxytyramín.
Poradie syntézy katecholamínov je pomerne jednoduché: tyrozín → dihydroxyfenylalanín (DOPA) → dopamín → norepinefrín → adrenalín. Tyrozín sa do tela dostáva s jedlom, ale môže sa tvoriť aj z fenylalanínu v pečeni pôsobením fenylalanínhydroxylázy. Konečné produkty premeny tyrozínu v tkanivách sú rôzne. V dreni nadobličiek proces pokračuje do štádia tvorby adrenalínu, v zakončeniach sympatických nervov - norepinefrínu, v niektorých neurónoch centrálneho nervového systému syntéza katecholamínov končí tvorbou dopamínu.
Premenu tyrozínu na DOPA katalyzuje tyrozínhydroxyláza, ktorej kofaktormi sú tetrahydrobiopterín a kyslík. Predpokladá sa, že tento enzým obmedzuje rýchlosť celého procesu biosyntézy katecholamínov a je inhibovaný konečnými produktmi procesu. Tyrozínhydroxyláza je hlavným cieľom regulačných účinkov na biosyntézu katecholamínov.
Premenu DOPA na dopamín katalyzuje enzým DOPA dekarboxyláza (kofaktor - pyridoxalfosfát), ktorý je relatívne nešpecifický a dekarboxyluje iné aromatické L-aminokyseliny. Existujú však náznaky možnosti modifikácie syntézy katecholamínov zmenou aktivity tohto enzýmu. Niektorým neurónom chýbajú enzýmy pre ďalšiu premenu dopamínu a ten je konečným produktom. Iné tkanivá obsahujú dopamín beta-hydroxylázu (kofaktory - meď, kyselina askorbová a kyslík), ktorá premieňa dopamín na norepinefrín. V dreni nadobličiek (ale nie v zakončeniach sympatických nervov) je prítomný fenyletanolamín - metyltransferáza, ktorá tvorí adrenalín z norepinefrínu. V tomto prípade slúži S-adenozylmetionín ako donor metylových skupín.
Je dôležité si uvedomiť, že syntéza fenyletanolamín-N-metyltransferázy je indukovaná glukokortikoidmi vstupujúcimi do drene z kôry cez portálny venózny systém. To môže vysvetľovať skutočnosť, že v jednom orgáne sú kombinované dve rôzne endokrinné žľazy. Význam glukokortikoidov pre syntézu adrenalínu je zdôraznený skutočnosťou, že bunky drene nadobličiek, ktoré produkujú noradrenalín, sa nachádzajú okolo arteriálnych ciev, zatiaľ čo bunky produkujúce adrenalín prijímajú krv prevažne z venóznych dutín nachádzajúcich sa v kôre nadobličiek.
K rozkladu katecholamínov dochádza hlavne pod vplyvom dvoch enzýmových systémov: katechol-O-metyltransferázy (COMT) a monoaminooxidázy (MAO). Hlavné dráhy rozkladu adrenalínu a noradrenalínu sú schematicky znázornené na obr. 54. Pod vplyvom COMT v prítomnosti donora metylovej skupiny S-adrenosylmetionínu sa katecholamíny premieňajú na normetanefrín a metanefrín (3-O-metylderiváty noradrenalínu a adrenalínu), ktoré sa pod vplyvom MAO premieňajú na aldehydy a potom (v prítomnosti aldehydoxidázy) na kyselinu vanilylmandľovú (VMA), hlavný produkt rozkladu noradrenalínu a adrenalínu. V rovnakom prípade, keď sú katecholamíny spočiatku vystavené MAO a nie COMT, premieňajú sa na 3,4-dioxomandľový aldehyd a potom pod vplyvom aldehydoxidázy a COMT na kyselinu 3,4-dioxomandľovú a VMC. V prítomnosti alkoholdehydrogenázy sa z katecholamínov môže tvoriť 3-metoxy-4-oxyfenylglykol, ktorý je hlavným konečným produktom degradácie adrenalínu a noradrenalínu v CNS.
Rozklad dopamínu je podobný, až na to, že jeho metabolitom chýba hydroxylová skupina na beta-uhlíkovom atóme, a preto namiesto kyseliny vanilylmandľovej vzniká kyselina homovanilová (HVA) alebo kyselina 3-metoxy-4-hydroxyfenyloctová.
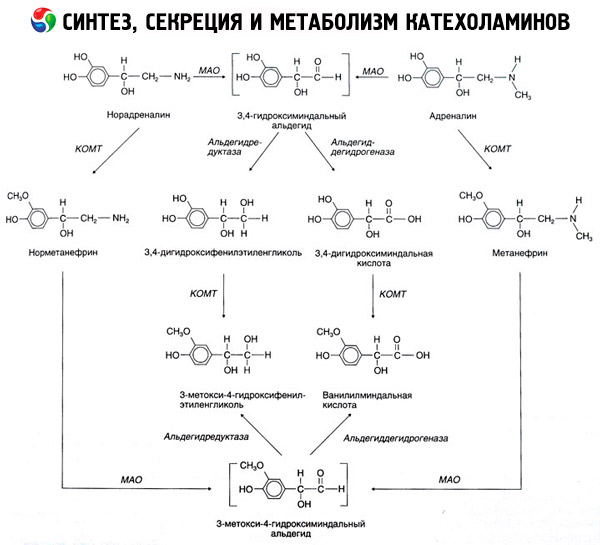
Predpokladá sa aj existencia chinoidnej dráhy oxidácie molekuly katecholamínu, ktorá môže produkovať medziprodukty s výraznou biologickou aktivitou.
Norepinefrín a adrenalín, ktoré vznikajú pôsobením cytozolických enzýmov v sympatických nervových zakončeniach a dreni nadobličiek, vstupujú do sekrečných granúl, ktoré ich chránia pred pôsobením degradačných enzýmov. Zachytenie katecholamínov granulami si vyžaduje výdaj energie. V chromafínnych granulách drene nadobličiek sú katecholamíny pevne viazané na ATP (v pomere 4:1) a špecifické proteíny - chromograníny, čo bráni difúzii hormónov z granúl do cytoplazmy.
Priamym stimulom pre sekréciu katecholamínov je zrejme prenikanie vápnika do bunky, stimulujúce exocytózu (fúzia membrány granúl s povrchom bunky a ich prasknutie s úplným uvoľnením rozpustného obsahu - katecholamínov, dopamínovej beta-hydroxylázy, ATP a chromogranínov - do extracelulárnej tekutiny).
Fyziologické účinky katecholamínov a ich mechanizmus účinku
Účinky katecholamínov začínajú interakciou so špecifickými receptormi cieľových buniek. Zatiaľ čo receptory pre hormóny štítnej žľazy a steroidné hormóny sú lokalizované vo vnútri buniek, receptory pre katecholamíny (ako aj pre acetylcholín a peptidové hormóny) sú prítomné na vonkajšom povrchu bunky.
Dlhodobo je známe, že v súvislosti s niektorými reakciami je adrenalín alebo noradrenalín účinnejší ako syntetický katecholamín izoproterenol, zatiaľ čo v súvislosti s inými je účinok izoproterenolu lepší ako účinky adrenalínu alebo noradrenalínu. Na tomto základe bola vyvinutá koncepcia o prítomnosti dvoch typov adrenoreceptorov v tkanivách: alfa a beta, pričom v niektorých z nich môže byť prítomný iba jeden z týchto dvoch typov. Izoproterenol je najsilnejším agonistom beta-adrenoreceptorov, zatiaľ čo syntetická zlúčenina fenylefrín je najsilnejším agonistom alfa-adrenoreceptorov. Prirodzené katecholamíny - adrenalín a noradrenalín - sú schopné interagovať s receptormi oboch typov, ale adrenalín vykazuje väčšiu afinitu k beta- a noradrenalín k alfa-receptorom.
Katecholamíny aktivujú srdcové beta-adrenergné receptory silnejšie ako beta-receptory hladkého svalstva, čo umožnilo rozdeliť beta typ na podtypy: beta1-receptory (srdce, tukové bunky) a beta2-receptory (priedušky, cievy atď.). Účinok izoproterenolu na beta1-receptory prevyšuje účinok adrenalínu a noradrenalínu iba 10-krát, zatiaľ čo na beta2-receptory pôsobí 100-1000-krát silnejšie ako prirodzené katecholamíny.
Použitie špecifických antagonistov (fentolamín a fenoxybenzamín pre alfa- a propranolol pre beta-receptory) potvrdilo primeranosť klasifikácie adrenoreceptorov. Dopamín je schopný interagovať s alfa- aj beta-receptormi, ale rôzne tkanivá (mozog, hypofýza, cievy) majú tiež svoje vlastné dopaminergné receptory, ktorých špecifickým blokátorom je haloperidol. Počet beta-receptorov sa pohybuje od 1000 do 2000 na bunku. Biologické účinky katecholamínov sprostredkované beta-receptormi sú zvyčajne spojené s aktiváciou adenylátcyklázy a zvýšením intracelulárneho obsahu cAMP. Hoci sú receptor a enzým funkčne prepojené, ide o rôzne makromolekuly. Guanozíntrifosfát (GTP) a ďalšie purínové nukleotidy sa pod vplyvom komplexu hormón-receptor podieľajú na modulácii aktivity adenylátcyklázy. Zvyšovaním aktivity enzýmu sa zdá, že znižujú afinitu beta receptorov k agonistom.
Fenomén zvýšenej citlivosti denervovaných štruktúr je už dlho známy. Naopak, dlhodobé pôsobenie agonistov znižuje citlivosť cieľových tkanív. Štúdium beta receptorov umožnilo vysvetliť tieto javy. Ukázalo sa, že dlhodobé pôsobenie izoproterenolu vedie k strate citlivosti adenylátcyklázy v dôsledku zníženia počtu beta receptorov.
Proces desenzibilizácie nevyžaduje aktiváciu syntézy bielkovín a je pravdepodobne spôsobený postupnou tvorbou ireverzibilných komplexov hormón-receptor. Naopak, zavedenie 6-oxidopamínu, ktorý ničí sympatické zakončenia, je sprevádzané zvýšením počtu reagujúcich beta-receptorov v tkanivách. Je možné, že zvýšenie aktivity sympatického nervového systému spôsobuje aj vekom podmienenú desenzibilizáciu ciev a tukového tkaniva vo vzťahu ku katecholamínom.
Počet adrenoreceptorov v rôznych orgánoch môže byť riadený inými hormónmi. Estradiol teda zvyšuje a progesterón znižuje počet alfa-adrenoreceptorov v maternici, čo je sprevádzané zodpovedajúcim zvýšením a znížením jej kontraktilnej odpovede na katecholamíny. Ak je intracelulárnym „druhým poslom“ tvoreným pôsobením agonistov beta-receptorov určite cAMP, potom je situácia s ohľadom na prenášač alfa-adrenergných účinkov komplikovanejšia. Predpokladá sa existencia rôznych mechanizmov: zníženie hladiny cAMP, zvýšenie obsahu cAMP, modulácia dynamiky bunkového vápnika atď.
Na reprodukciu rôznych účinkov v tele sú zvyčajne potrebné dávky adrenalínu, ktoré sú 5 až 10-krát menšie ako noradrenalín. Hoci je noradrenalín účinnejší vo vzťahu k a- a beta1-adrenoreceptorom, je dôležité mať na pamäti, že oba endogénne katecholamíny sú schopné interagovať s alfa- aj beta-receptormi. Preto biologická odpoveď daného orgánu na adrenergnú aktiváciu do značnej miery závisí od typu receptorov, ktoré sú v ňom prítomné. To však neznamená, že selektívna aktivácia nervového alebo humorálneho spoja sympaticko-nadobličkového systému je nemožná. Vo väčšine prípadov sa pozoruje zvýšená aktivita jeho rôznych spojov. Všeobecne sa teda uznáva, že hypoglykémia reflexne aktivuje dreň nadobličiek, zatiaľ čo pokles krvného tlaku (posturálna hypotenzia) je sprevádzaný najmä uvoľňovaním noradrenalínu zo zakončení sympatických nervov.
Adrenoreceptory a účinky ich aktivácie v rôznych tkanivách
Systém, orgán |
Typ adrenergného receptora |
Reakcia |
Kardiovaskulárny systém: |
||
Srdce |
Beta verzia |
Zvýšená srdcová frekvencia, vodivosť a kontraktilita |
Arterioly: |
||
Koža a sliznice |
Alfa |
Zníženie |
Kostrové svaly |
Beta verzia |
Expanzia Kontrakcia |
Brušné orgány |
Alfa (viac) |
Zníženie |
Beta verzia |
Rozšírenie |
|
Žily |
Alfa |
Zníženie |
Dýchací systém: |
||
Svaly priedušiek |
Beta verzia |
Rozšírenie |
Tráviaci systém: |
||
Žalúdok |
Beta verzia |
Znížené motorické zručnosti |
Črevá |
Alfa |
Kontrakcia zvieračov |
Slezina |
Alfa |
Zníženie |
Beta verzia |
Relaxácia |
|
Exokrinný pankreas |
Alfa |
Znížená sekrécia |
Urogenitálny systém: |
Alfa |
Kontrakcia zvierača |
Močový mechúr |
Beta verzia |
Relaxácia ejektorového svalu |
Mužské genitálie |
Alfa |
Ejakulácia |
Oči |
Alfa |
Rozšírenie zreníc |
Koža |
Alfa |
Zvýšené potenie |
Slinné žľazy |
Alfa |
Vylučovanie draslíka a vody |
Beta verzia |
Sekrécia amylázy |
|
Endokrinné žľazy: |
||
Ostrovčeky pankreasu |
||
Beta bunky |
Alfa (viac) |
Znížená sekrécia inzulínu |
Beta verzia |
Zvýšená sekrécia inzulínu |
|
Alfa bunky |
Beta verzia |
Zvýšená sekrécia glukagónu |
8-článková |
Beta verzia |
Zvýšená sekrécia somatostatínu |
Hypotalamus a hypofýza: |
||
Somatotrofy |
Alfa |
Zvýšená sekrécia STH |
Beta verzia |
Znížená sekrécia STH |
|
Laktotrofy |
Alfa |
Znížená sekrécia prolaktínu |
Tyreotrofi |
Alfa |
Znížená sekrécia TSH |
Kortikotrofy |
Alfa |
Zvýšená sekrécia ACTH |
| beta | Znížená sekrécia ACTH | |
Štítna žľaza: |
||
Folikulárne bunky |
Alfa |
Znížená sekrécia tyroxínu |
Beta verzia |
Zvýšená sekrécia tyroxínu |
|
Parafolikulárne (K) bunky |
Beta verzia |
Zvýšená sekrécia kalcitonínu |
Prištítne telieska |
Beta verzia |
Zvýšená sekrécia PTH |
Obličky |
Beta verzia |
Zvýšená sekrécia renínu |
Žalúdok |
Beta verzia |
Zvýšená sekrécia gastrínu |
BX |
Beta verzia |
Zvýšená spotreba kyslíka |
Pečeň |
? |
Zvýšená glykogenolýza a glukoneogenéza s uvoľňovaním glukózy; zvýšená ketogenéza s uvoľňovaním ketónových teliesok |
Tukové tkanivo |
Beta verzia |
Zvýšená lipolýza s uvoľňovaním voľných mastných kyselín a glycerolu |
Kostrové svaly |
Beta verzia |
Zvýšená glykolýza s uvoľňovaním pyruvátu a laktátu; znížená proteolýza so zníženým uvoľňovaním alanínu a glutamínu |
Je dôležité vziať do úvahy, že výsledky intravenózneho podávania katecholamínov nie vždy adekvátne odrážajú účinky endogénnych zlúčenín. To platí najmä pre norepinefrín, pretože v tele sa uvoľňuje prevažne nie do krvi, ale priamo do synaptických štrbín. Preto endogénny norepinefrín aktivuje napríklad nielen vaskulárne alfa receptory (zvýšený krvný tlak), ale aj beta receptory srdca (zvýšená srdcová frekvencia), zatiaľ čo podanie norepinefrínu zvonku vedie najmä k aktivácii vaskulárnych alfa receptorov a reflexnému (cez vagus) spomaleniu srdcového tepu.
Nízke dávky adrenalínu aktivujú prevažne beta receptory svalových ciev a srdca, čo vedie k zníženiu periférneho cievneho odporu a zvýšeniu srdcového výdaja. V niektorých prípadoch môže prevládať prvý účinok a po podaní adrenalínu sa vyvinie hypotenzia. Vo vyšších dávkach adrenalín aktivuje aj alfa receptory, čo je sprevádzané zvýšením periférneho cievneho odporu a na pozadí zvýšenia srdcového výdaja vedie k zvýšeniu arteriálneho tlaku. Jeho účinok na cievne beta receptory je však tiež zachovaný. V dôsledku toho zvýšenie systolického tlaku prevyšuje podobný ukazovateľ diastolického tlaku (zvýšenie pulzného tlaku). Pri podávaní ešte vyšších dávok začínajú prevládať alfa-mimetické účinky adrenalínu: systolický a diastolický tlak sa zvyšujú paralelne, ako pod vplyvom norepinefrínu.
Účinok katecholamínov na metabolizmus pozostáva z ich priamych a nepriamych účinkov. Prvé sa realizujú hlavne prostredníctvom beta receptorov. Zložitejšie procesy sú spojené s pečeňou. Hoci sa zvýšená glykogenolýza pečene tradične považuje za výsledok aktivácie beta receptorov, existujú aj dôkazy o zapojení alfa receptorov. Nepriame účinky katecholamínov sú spojené s moduláciou sekrécie mnohých ďalších hormónov, ako je inzulín. V účinku adrenalínu na jeho sekréciu jednoznačne prevláda alfa adrenergná zložka, pretože sa preukázalo, že akýkoľvek stres je sprevádzaný inhibíciou sekrécie inzulínu.
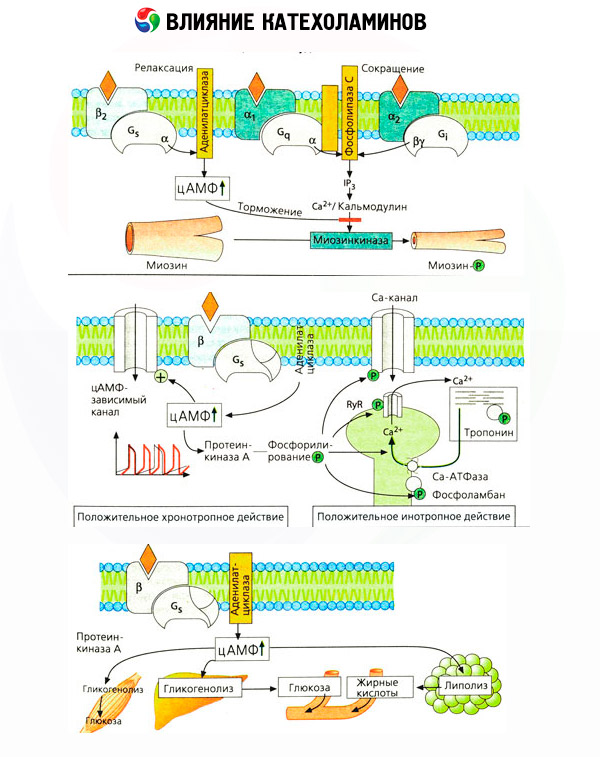
Kombinácia priamych a nepriamych účinkov katecholamínov spôsobuje hyperglykémiu, spojenú nielen so zvýšenou produkciou glukózy v pečeni, ale aj s inhibíciou jej využitia periférnymi tkanivami. Zrýchlenie lipolýzy spôsobuje hyperlipacidémiu so zvýšeným dodávaním mastných kyselín do pečene a zintenzívnením produkcie ketónových teliesok. Zvýšená glykolýza vo svaloch vedie k zvýšenému uvoľňovaniu laktátu a pyruvátu do krvi, ktoré spolu s glycerolom uvoľneným z tukového tkaniva slúžia ako prekurzory pečeňovej glukoneogenézy.
Regulácia sekrécie katecholamínov. Podobnosť produktov a metód reakcie sympatického nervového systému a drene nadobličiek bola základom pre spojenie týchto štruktúr do jediného sympaticko-nadobličkového systému tela s rozdelením jeho nervových a hormonálnych väzieb. Rôzne aferentné signály sú koncentrované v hypotalame a centrách miechy a predĺženej miechy, odkiaľ pochádzajú eferentné správy, ktoré prechádzajú do bunkových telies pregangliových neurónov nachádzajúcich sa v bočných rohoch miechy na úrovni VIII krčných - II-III bedrových segmentov.
Pregangliové axóny týchto buniek opúšťajú miechu a vytvárajú synaptické spojenia s neurónmi nachádzajúcimi sa v gangliách sympatického reťazca alebo s bunkami drene nadobličiek. Tieto pregangliové vlákna sú cholinergné. Prvý zásadný rozdiel medzi sympatickými postgangliovými neurónmi a chromafínnymi bunkami drene nadobličiek spočíva v tom, že tie druhé prenášajú cholinergný signál, ktorý prijímajú, nie nervovým vedením (postgangliové adrenergné nervy), ale humorálnou cestou, čím uvoľňujú adrenergné zlúčeniny do krvi. Druhý rozdiel spočíva v tom, že postgangliové nervy produkujú norepinefrín, zatiaľ čo bunky drene nadobličiek produkujú hlavne adrenalín. Tieto dve látky majú odlišné účinky na tkanivá.