
Hypofýza
Posledná kontrola: 23.04.2024

Všetok obsah iLive je lekársky kontrolovaný alebo kontrolovaný, aby sa zabezpečila čo najväčšia presnosť faktov.
Máme prísne smernice týkajúce sa získavania zdrojov a len odkaz na seriózne mediálne stránky, akademické výskumné inštitúcie a vždy, keď je to možné, na lekársky partnerské štúdie. Všimnite si, že čísla v zátvorkách ([1], [2] atď.) Sú odkazmi na kliknutia na tieto štúdie.
Ak máte pocit, že niektorý z našich obsahov je nepresný, neaktuálny alebo inak sporný, vyberte ho a stlačte kláves Ctrl + Enter.
Hypofýzy (hypofýza, s.glandula pituitaria) uložené v podmozgovej fossa Sella sphenoid kostí a lebečnej dutiny je oddelená od pevnej prívesok mozgovej membrány, tvoriace membránové sedlo. Prostredníctvom otvoru v tejto diafragme je hypofýza pripojená k lieviku hypotalamu stredného mozgu. Prierezová veľkosť hypofýzy je 10-17 mm, anteroposterior - 5-15 mm, vertikálne - 5-10 mm. Hmotnosť hypofýzy u mužov je približne 0,5 g, u žien je to 0,6 g. Vonku je hypofýza pokrytá kapsulou.
V súlade s vývojom hypofýzy z dvoch rôznych mikróbov v tele rozlíšiť dve časti - prednú a zadnú stranu. Adenohypofýzy alebo predného laloku (adenohypofýzy, s.lobus predné), tým väčšia je 70 až 80% z celkovej hmotnosti hypofýzy. Je hustší ako zadný lalok. V prednom lalokom distálnej časti sa izoluje (Pars distalis), ktorý zaberá na prednú časť hypofýzy fossa, stredná časť (pars intermedia), usporiadaný na rozhraní zadného podielu a bugornuyu časť (pars tuberalis), pričom a pripojená k lieviku hypotalamu. Vzhľadom na množstvo ciev predného laloku má svetlo žltú farbu s červenkastým zafarbením. Pletivá adenohypofýzy glandulární zástupcovia niekoľkých typov buniek, ktoré sú umiestnené medzi prameňmi sínusové kapiláry. Polovica (50%) z adenohypofýzy buniek sú hromafilnymi adenocytes, ktoré majú vo svojich cytoplazmatických jemné granule, dobre zafarbených soľou chrómu. Tento acidofilné adenocytes (40% z adenohypofýzy buniek) a bazofilné adenocytes {10%). Počet bazofilných adenocytes zahŕňajú gonadotropné, kortikotropnye thyreotropní endocrinocytes. Chromofobní adenocytes malé, majú veľké jadro a malé množstvo cytoplazmy. Tieto bunky sa považujú za prekurzory chromofilných adenocytov. Ďalšími 50% buniek adenohypofýzy sú chromofóbne adenocyty.
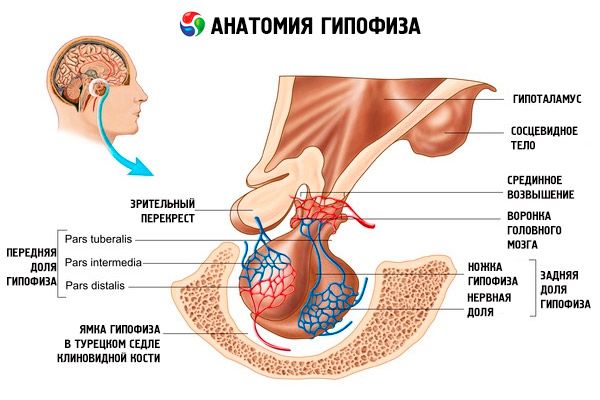
Neurohypofýza alebo zadné lalok (neurohypofýza, s.lobus zadné), skladajúci sa z nervového frakcie (lobus nervosus), ktorý je umiestnený v zadnej časti hypofýzy jamky a lievikom (infundibulum), ktorá sa nachádza za bugornoy časť adenohypofýzy. Hypofýzy lalôčik tvorený gliových buniek (bunky hypofýzy), nervové vlákna, ktorý je vedený od neurosecretory jadrách hypotalamu v neurohypofýzy a neurosekrečních buniek.
Hypofýza pomocou nervových vlákien (cestičiek) a krvných ciev je funkčne spojená s hypotalamom prechodného mozgu, ktorý reguluje činnosť hypofýzy. Hypofýza a hypotalamus, spolu s ich neuroendokrinnými, vaskulárnymi a nervovými spojeniami, sa zvyčajne považujú za systém hypofýzy-hypofýzy.
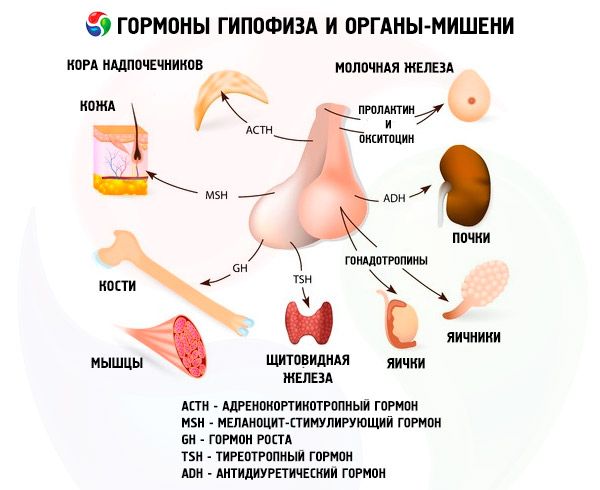
Hormóny predné a hypofýzy ovplyvniť veľa telesných funkcií, najmä prostredníctvom ďalších žliaz s vnútornou sekréciou. Vo predného laloku hypofýzy acidofilných adenocytes (alfa) bunky produkujú somotropny hormón (HGH), ktoré sa zúčastňujú regulácie rastu a vývoja mladého organizmu. Kortikotropnye endocrinocytes vylučujú kortikotropín (ACTH), ktorý stimuluje sekréciu steroidných hormónov nadobličiek. Tirotropnye endocrinocytes vylučujú tirotropny hormón (TSH), ktorý ovplyvňuje vývoj štítnej žľazy a aktivácia produkciu jej hormónov. Gonadotropným hormóny: folikuly stimulujúci hormón (FSH), luteinizačného hormónu (LH) a prolaktínu - vplyv puberta telo, reguláciu a stimuláciu vývoja folikulov vo vaječníku, ovulácie, prsia rast a produkciu mlieka u žien, proces spermatogenézy u mužov. Tieto hormóny sú produkované bazofilnými adenocytmi beta-buniek ). Tu lipotropic faktory vylučované z hypofýzy, ktoré majú vplyv na mobilizáciu a využitie tuku v tele. Medziľahlá časť predného laloku vytvorený hormónu stimulujúceho melanocyty, ktorý riadi tvorbu pigmentov - melanín - v tele.
Neurosekrečné bunky superoptických a paraventrikulárnych jadier v hypotalame produkujú vazopresín a oxytocín. Tieto hormóny sa prenášajú do buniek zadného laloku hypofýzy pozdĺž axónov, ktoré tvoria hypotalamo-hypofyzárny trakt. Z zadného laloku hypofýzy sa tieto látky dostávajú do krvi. Hormón vazopresínu má vazokonstrikčný a antidiuretický účinok, na ktorý sa tiež nazýva antidiuretický hormón (ADH). Oxytocín má stimulačný účinok na kontraktilitu svalov maternice, zvyšuje mlieko z mliečnej žľazy dojčiace, inhibuje funkciu luteum vývoja a corpus, má vplyv na hladké zmeny tónových (neischerchennyh) svaly gastrointestinálneho traktu.
Vývoj hypofýzy
Predná časť hypofýzy sa vyvíja z epitelu dorzálnej steny ústnej dutiny vo forme prstencovitého výrastku (Rathkeho vrecko). Tento ektodermálny výčnelok rastie smerom k dnu budúcej III. Komory. K nemu od spodnej povrch druhej bublinkové mozgu (budúce spodnej komory III) vyrážať rastie, z ktorej vytvoril šedej hrbolček lievikom a zadný lalok hypofýzy.
 [1], [2], [3], [4], [5], [6], [7], [8], [9], [10], [11], [12], [13]
[1], [2], [3], [4], [5], [6], [7], [8], [9], [10], [11], [12], [13]
Plavidlá a nervy hypofýzy
Horné a dolné hypofýzové tepny sú nasmerované z vnútorných krčných ciev a krvných ciev mozgovej arteriálnej kružnice k hypofýze. Horná hypofýzy tepna ísť do šedej jadro lievikom a hypotalamu anastomose medzi sebou a tu tvorí preniká do mozgového tkaniva kapilár - gemokapillyarnuyu primárnej siete. Z dlhých a krátkych slučiek tejto siete sa vytvárajú portálne žily, ktoré smerujú k prednému laloku hypofýzy. V parenchýme predného laloku hypofýzy sa tieto žily disociujú do širokých sínusových kapilár, ktoré tvoria sekundárnu hemokapilárnu sieť. Zadný lalok hypofýzy je prevažne krv tečúcou cez spodnú hypofýzu. Medzi hornou a dolnou hypofýzou existujú dlhé arteriálne anastomózy. Odtok žilovej krvi zo sekundárnej hemokapilárnej siete sa uskutočňuje systémom žíl, ktoré prúdia do kavernóznych a medzigitálnych dutín tvrdého mozgu.
Inervácia hypofýzy zahŕňa sympatické vlákna, ktoré prenikajú do orgánu spolu s tepnami. Postgangliové sympatické nervové vlákna sa pohybujú od prelínania vnútornej krčnej tepny. Okrem toho sa v zadnom laloku hypofýzy objavujú mnohé výrastky procesov neurosekrečných buniek nachádzajúcich sa v jadre hypotalamu.
Vekové charakteristiky hypofýzy
Priemerná hmotnosť hypofýzy u novorodencov dosahuje 0,12 g. Telesná hmotnosť sa zdvojnásobí na 10 a trojnásobne sa zvyšuje o 15 rokov. Do veku 20 rokov dosahuje hmotnosť hypofýzy maximálne (530-560 mg) a v nasledujúcich vekových kategóriách sa takmer nemení. Po 60 rokoch dochádza k miernemu poklesu hmotnosti tejto endokrinnej žľazy.
[14], [15], [16], [17], [18], [19], [20], [21], [22], [23], [24], [25], [26]
Hormóny hypofýzy
Jednota nervovej a hormonálnej regulácie v tele je zabezpečená tesným anatomickým a funkčným spojením hypofýzy a hypotalamu. Tento komplex určuje stav a fungovanie celého endokrinného systému.
Hlavné endokrinné žľazy, ktorá produkuje rad peptidových hormónov, ktoré priamo regulujú funkciu periférnych žliaz - hypofýzu. Je červeno-šedý tvare fazuľa útvar, na ktoré sa vzťahuje vláknitého kapsule s hmotnosťou 0,5-0,6 g sa mierne líšia v závislosti od pohlavia a veku osoby. Je všeobecne známe, rozdelenie hypofýzy do dvoch častí, rôzne vývoja, štruktúry a funkcie: predné distálnej - adenohypofýzy a späť - neurohypofýzy. Prvé asi 70% z celkovej hmotnosti prostaty a je rozdelený na distálnej, Voronkov a medziľahlé časti, druhý - na zadnej časti, alebo frakcie, a hypofýzy stonky. Žľaza sa nachádza v podmozgovej fossa sella klinová kosť a cez nohu je pripojený do mozgu. Horná časť predného laloku je pokrytá vizuálnym prekrížením a vizuálnymi cestami. Perfúzie hypofýzy veľmi hojné vetvy a sa uskutočňuje z vnútornej krčnej tepny (horná a dolná hypofýzy tepien) a konáre mozgovej tepny kruhu. Horná hypofýzy tepna prekrvenie podieľa adenohypofýzy a nižšie - neurohypofýzy, kde pri kontakte s neurosecretory axónov koncovkami hypotalamus magnocellular. Prvá časť strednej reputáciou hypotalamu, ktoré sú rozptýlené v kapilárnej siete (primárna kapilárnej plexus). Tieto kapiláry (ktoré kontaktné terminály axónov malé mediobasal hypotalamu neurosekrečních buniek) zhromaždených v portálnej žile Descending pozdĺž nohy v hypofýze adenohypofýzy pásoch, kde opäť rozdelené do sínusovej kapilár reťazca (sekundárne kapilárnej plexu). Tak, krv, vopred preosiate cez stredné reputáciou hypotalamu, kde adenogipofizotropnymi obohatené hypotalamus hormón (kortikotropín uvoľňujúci hormón), dostáva do adenohypofýzy.
Odtok krvi, nasýtené adenogipofizarnymi hormóny početných kapilár sekundárne plexus žily sa vykonáva pomocou systému, ktorý zase prúdi do žilového dutín v dura mater a do krvného obehu. Tak, hypofýza portál systém k smeru zostupného toku krvi z hypotalamu je súčasťou zložitého mechanizmu morphofunctional neurohumorální tropný kontrolné funkcie adenohypofýzy.
Inervácia hypofýzy sa uskutočňuje sympatickými vláknami, ktoré sledujú hypofýzové tepny. Začínajú im postgangliové vlákna, prechádzajúce cez vnútorný karotidový plexus, spojené s hornými cervikálnymi uzlinami. Neexistuje žiadna priama inervácia adenohypofýzy z hypotalamu. Nervové vlákna neurosecretorových jadier hypotalamu vstupujú do zadného laloku.
Adenohypofýza v histologickej architektonike je veľmi zložitá forma. Rozlišuje dva typy glandulárnych buniek - chromofóbne a chromofilné. Tie sú ďalej rozdelené do eozinofilov a bazofilov (hypofýzy histologicky popis je uvedený v príslušnej časti manuálu). Malo by však byť poznamenané, že hormóny produkoval glandulární bunky, ktoré tvoria parenchým adenohypofýzy, vzhľadom k nedávnemu rôznorodosti v trochu odlišné v ich chemickej povahe a jemnú štruktúru sekretiziruyuschih buniek by mala zodpovedať biosyntézy vlastnosti každého z nich. Ale niekedy v adenohypofýze je možné pozorovať prechodné formy žľazových buniek, ktoré sú schopné produkovať niekoľko hormónov. Existuje dôkaz, že množstvo glandulárnych buniek adenohypofýzy nie je vždy určené geneticky.
Pod membránou tureckého sedla je lieviková časť predného laloku. Pokrýva pätu hypofýzy a kontaktuje sivý návrší. Táto časť adenohypofýzy sa vyznačuje prítomnosťou epiteliálnych buniek a bohatým zásobovaním krvou. Je tiež hormonálne aktívny.
Stredná (stredná) časť hypofýzy pozostáva z niekoľkých vrstiev veľkých sekretárne aktívnych bazofilných buniek.
Hypofýza prostredníctvom svojich hormónov nesie rôzne funkcie. Vo svojej prednej laloku vyrobeného adrenokortikotropinu (ACTH), thyreotropní (TSH), folikuly stimulujúci hormón (FSH), luteinizačný hormón (LH), lipotropic hormónu a rastový hormón. - Somatotropic (SRT a prolaktínu v medziľahlej laloku syntetizovanej melanocyty stimulujúci hormón (MSH), a V zadnej časti sa nahromadia vazopresín a oxytocín.
AktG
Hypofýzové hormóny predstavujú skupinu proteínových a peptidových hormónov a glykoproteínov. Z hormónov predného laloku hypofýzy je ACTH najviac študovaný. Vyrába sa z bazofilných buniek. Jeho hlavnou fyziologickou funkciou je stimulácia biosyntézy a sekrécie steroidných hormónov kôrou nadobličiek. ACTH tiež vykazuje stimuláciu melanocytov a lipotropnú aktivitu. V roku 1953 bola izolovaná vo svojej čistej forme. Neskôr sa vytvorila jej chemická štruktúra, pozostávajúca z 39 aminokyselinových zvyškov u človeka a množstva cicavcov. ACTH nemá špecifickú špecifickosť. V súčasnej dobe sa uskutočňuje chemická syntéza samotného hormónu a rôznych účinnejších ako prírodných hormónov, fragmentov jeho molekuly. V štruktúre hormónu dve časti peptidového reťazca, z ktorých jedna poskytuje detekciu a väzbu ACTH na receptor a druhá - poskytuje biologický účinok. S receptorom ACTH sa zdá, že sa viaže v dôsledku interakcie elektrických nábojov s hormónom a receptorom. Úloha biologického efektora ACTH uskutočňuje fragment molekuly 4-10 (Met-Glu-Gis-Fen-Arg-Tri-Tri).
Melanocyt-stimulujúca aktivita ACTH je spôsobená prítomnosťou N-koncovej oblasti v molekule pozostávajúcej z 13 aminokyselinových zvyškov a opakovaním štruktúry hormónu stimulujúceho alfa-melanocyty. Rovnaké miesto obsahuje heptapeptid, ktorý je prítomný v iných hormónoch hypofýzy a má niektoré adrenokortikotropné, stimulujúce melanocyty a lipotropné aktivity.
Kľúčovým bodom pôsobenia ACTH je aktivácia enzýmu proteínkinázy v cytoplazme za účasti cAMP. Fosforylovaný proteín kináza aktivuje enzým esterázy prevedie esterov cholesterolu na voľné mastné látky v kvapôčkach. Proteín sa syntetizuje v cytoplazme ako dôsledok fosforylácie ribozóme väzbového voľný cholesterol stimuluje cytochróm P-450, a prenášať ich z lipidových kvapiek v mitochondriách, kde sú všetky enzýmy, ktoré zaisťujú premenu cholesterolu na kortikosteroidy.
[27], [28], [29], [30], [31], [32], [33], [34], [35], [36], [37], [38], [39]
Tyrotropický hormón
TSH - tyreotropín - hlavný regulátor vývoja a fungovania štítnej žľazy, procesy syntézy a sekrécie hormónov štítnej žľazy. Tento komplexný proteín - glykoproteín - pozostáva z alfa a beta podjednotiek. Štruktúra prvej podjednotky sa zhoduje s alfa podjednotkou luteinizačného hormónu. Okrem toho sa do značnej miery zhoduje s rôznymi druhmi zvierat. Sekvencia aminokyselinových zvyškov v ľudskej beta-podjednotke ľudského TSH sa dešifruje a skladá sa z 119 aminokyselinových zvyškov. Je možné poznamenať, že beta podjednotky ľudského TSH a hovädzieho dobytka sú v mnohých ohľadoch podobné. Biologické vlastnosti a charakter biologickej aktivity glykoproteínových hormónov sa stanovujú pomocou beta podjednotky. Zabezpečuje tiež interakciu hormónu s receptormi v rôznych cieľových orgánoch. Avšak beta podjednotka vo väčšine zvierat vykazuje špecifickú aktivitu až po jej spojení s alfa-podjednotkou, ktorá pôsobí ako druh aktivátora hormónu. Posledne uvedená s rovnakou pravdepodobnosťou indukuje luteinizačné, folikuly stimulujúce a tyrotropické aktivity, ktoré sú určené vlastnosťami beta podjednotky. Zistená podobnosť nám umožňuje dospieť k záveru, že tieto hormóny pochádzajú z vývoja z jedného spoločného prekurzora, beta podjednotka určuje imunologické vlastnosti hormónov. Predpokladá sa, že alfa podjednotka chráni beta podjednotku pred pôsobením proteolytických enzýmov a tiež uľahčuje jej transport z hypofýzy do periférnych cieľových orgánov.
Gonadotropné hormóny
Gonadotropíny sú prítomné v tele v podobe LH a FSH. Funkčný účel týchto hormónov zvyčajne obmedzuje na poskytovanie reprodukčných procesov u jedincov oboch pohlaví. Tak ako TTG sú komplexné proteíny - glykoproteíny. FSH indukuje maturáciu folikulov vo vaječníkoch u žien a stimuluje spermatogenézu u mužov. LH spôsobuje u žien prasknutie folikulu s tvorbou žltého tela a stimuluje sekréciu estrogénov a progesterónu. U mužov tento istý hormón urýchľuje vývoj intersticiálneho tkaniva a sekrécie androgénov. Účinky gonadotropínov sú navzájom závislé a pokračujú synchrónne.
Dynamika sekrécie gonadotropínu u žien sa mení počas menštruačného cyklu a je študovaná dostatočne podrobne. V predovulovacej (folikulárnej) fáze cyklu je obsah LH na pomerne nízkej úrovni a FSH sa zvyšuje. Vzhľadom k tomu, dozrievanie folikulov sekrécie estradiolu zvýšené, čím sa zvyšuje produkcia gonadotropínov hypofýzou a výskytom cyklov ako LH a FSH a pod. E., sexuálne steroidy stimulujú sekréciu gonadotropínov.
V súčasnosti sa určuje štruktúra LH. Rovnako ako TTG, pozostáva z 2 podjednotiek: a a p. Štruktúra alfa podjednotky LH u rôznych druhov zvierat sa z veľkej časti zhoduje, zodpovedá štruktúre alfa-podjednotky TSH.
Štruktúra beta-podjednotke LH sa výrazne líši od štruktúry TSH beta podjednotky, aj keď má štyri rovnaké časti peptidového reťazca zložené z 4-5 aminokyselinových zvyškov. TTG, že sú lokalizované v polohách 27 až 31, 51 až 54, 65-68 a 78-83. Vzhľadom k tomu, beta podjednotka LH a TSH určuje špecifickú biologickú aktivitu hormónov, možno predpokladať, že homológne oblasti v štruktúre LH a TSH by mala poskytnúť beta-podjednotku alfa-podjednotku a štruktúrou líšia plochách - zodpovedné za špecificitu biologickej aktivity hormónu.
Natívne LH je veľmi stabilný voči pôsobeniu proteolytických enzýmov, ale beta podjednotku sa rýchlo štiepi chymotrypsínu, a tvrdý-podjednotka sa hydrolyzuje enzýmom, tj. E. To má ochrannú úlohu, zabránenie prístupu k chymotrypsínu peptidových väzieb.
Pokiaľ ide o chemickú štruktúru FSH, v súčasnosti výskumníci nedostali konečné výsledky. Rovnako ako LH, FSH pozostáva z dvoch podjednotiek, ale beta-podjednotka FSH sa líši od beta-podjednotky LH.
Prolaktín
V procese reprodukcie sa aktívne zúčastňuje ďalší hormón, prolaktín (laktogénny hormón). Hlavné fyziologické vlastnosti prolaktínu u cicavcov sa prejavujú vo forme stimulácie vývoja mliečnych žliaz a laktácie, rastu mazových žliaz a vnútorných orgánov. Podporuje účinok steroidov na sekundárne sexuálne vlastnosti u mužov, stimuluje sekrečnú aktivitu žltého tela u myší a potkanov a podieľa sa na regulácii metabolizmu tukov. Veľa pozornosti sa venuje prolaktínu v posledných rokoch ako regulátor správania matky, táto polyfunkčnosť sa vysvetľuje jeho evolučným vývojom. Je to jeden z dávnych hormónov hypofýzy a nachádza sa aj v obojživelníkov. V súčasnosti sa štruktúra prolaktínu niektorých cicavčích druhov úplne dešifrovala. Avšak nedávno vedci vyjadrili pochybnosti o existencii takéhoto hormónu u ľudí. Mnohí verili, že jeho funkcia je vykonávaná rastovým hormónom. Teraz máme presvedčivé dôkazy o prítomnosti prolaktínu u ľudí a čiastočne rozlíšili jeho štruktúru. Prolaktínové receptory aktívne viažu rastový hormón a placentárny laktogén, čo naznačuje jediný mechanizmus pôsobenia troch hormónov.
Somatotropínu
Ešte širšie spektrum účinku ako prolaktín má rastový hormón - somatotropín. Rovnako ako prolaktín, produkuje aj acidofilné bunky adenohypofýzy. STG stimuluje rast skeletu, aktivuje biosyntézu proteínu, poskytuje efekt mobilizujúci tuky, podporuje zvýšenie telesnej hmotnosti. Okrem toho koordinuje výmenné procesy.
Účasť hormónu v tomto druhu je potvrdená skutočnosťou, že sa výrazne zvýši sekrécia hypofýzy, napríklad poklesom obsahu cukru v krvi.
Chemická štruktúra tohto ľudského hormónu je teraz plne zavedená - 191 aminokyselinových zvyškov. Jeho primárna štruktúra je podobná štruktúre chorionického somatomamotropínu alebo placentárneho laktogénu. Tieto údaje naznačujú významnú evolučnú blízkosť dvoch hormónov, aj keď vykazujú rozdiely v biologickej aktivite.
Je potrebné zdôrazniť vysokú špecifickú špecifickosť daného hormónu - napríklad STH živočíšneho pôvodu je u ľudí neaktívny. Je to spôsobené reakciou medzi ľudskými a zvieracími receptormi STH a štruktúrou samotného hormónu. V súčasnej dobe sú prebiehajúce štúdie na identifikáciu aktívnych miest v komplexnej štruktúre STH, ktoré vykazujú biologickú aktivitu. Študujeme jednotlivé fragmenty molekuly, ktoré vykazujú iné vlastnosti. Napríklad po hydrolýze ľudského STH s pepsínom bol izolovaný peptid obsahujúci 14 aminokyselinových zvyškov a zodpovedajúci oblasti molekuly 31-44. Nemal účinok rastu, ale lipotropná aktivita bola významne lepšia ako pôvodný hormón. Ľudský rastový hormón, na rozdiel od podobného hormónu u zvierat, má významnú laktogénnu aktivitu.
V mnohých adenohypofýzy syntetizované ako peptidové a proteínové látky s obsahom tuku-mobilizačné účinnosť a tropný hormóny hypofýzy - ACTH, rastového hormónu, TSH a ďalšie - majú lipotropic účinok. V posledných rokoch boli vyčlenené najmä beta- a y-lipotropné hormóny (LPG). Medzi najviac skúmaný Biologické vlastnosti beta-LPG, ktorý, okrem lipotropic aktivita bola tiež melanocyty, kortikotropinstimuliruyuschee a hypocalcemic účinok a dáva účinok inzulínu.
V súčasnosti sa dešifruje primárna štruktúra LPG oviec (90 aminokyselinových zvyškov), lipotropných hormónov ošípaných a hovädzieho dobytka. Tento hormón má špecifickú špecifickosť, hoci štruktúra centrálnej časti beta-LPG u rôznych druhov je rovnaká. Určuje biologické vlastnosti hormónu. Jeden z fragmentov tohto miesta sa nachádza v štruktúre alfa-MSH, beta-MSH, ACTH a beta-LPG. Predpokladá sa, že tieto hormóny pochádzajú z toho istého prekurzora v priebehu vývoja. Y-LPG má slabšiu lipotropnú aktivitu ako beta-LPG.
Melanocyty stimulujúci hormón
Tento hormón je syntetizovaný v medziľahlé laloku hypofýzy, na jeho biologické funkcie stimuláciu pokožky pigment biosyntézy melanínu, zvyšuje veľkosť a množstvo pigmentových melanocytov v kožné bunky obojživelníkov. Tieto vlastnosti MSH sa používajú pri biologickom testovaní hormónu. Existujú dva typy hormónov: alfa a beta-MSH. Ukazuje sa, že alfa-MSH nemá špecifickú špecifickosť a má rovnakú chemickú štruktúru u všetkých cicavcov. Jeho molekulou je peptidový reťazec pozostávajúci z 13 aminokyselinových zvyškov. Beta-MSH má naopak špecifickú špecifickosť a jeho štruktúra sa líši u rôznych zvierat. Vo väčšine cicavcov sa ß-MSH molekula skladá z 18 aminokyselinových zvyškov a len u ľudí je predĺžená od konca aminoskupiny k štyrom aminokyselinovým zvyškom. Treba poznamenať, že alfa-MSH má určitú adrenokortikotropnú aktivitu a jeho účinok na správanie zvierat a ľudí je teraz preukázaný.
Oxytocín a vazopresín
Zadné lalok hypofýzy hromadí vazopresínu a oxytocínu, ktoré sú syntetizované v hypotalame: vazopresínu - v neurónoch supraoptic a oxytocín - paraventrikulyatornogo. Potom sa prenesú do hypofýzy. Treba zdôrazniť, že v hypotalame je prvý syntetizovaný prekurzor hormónu vazopresínu. Súčasne sa produkuje neurofizínový proteín prvého a druhého typu. Prvý sa viaže na oxytocín a druhý na vazopresín. Tieto komplexy migrujú ako neurosekreční granule v cytoplazme pozdĺž axónu a dosiahne zadné hypofýzy, kde nervové vlákna končia na cievne steny a granúl obsahu do krvi. Vasopresín a oxytocín sú prvými hormónmi hypofýzy s plne zavedenou aminokyselinovou sekvenciou. V ich chemickej štruktúre ide o nonapeptidy s jedným disulfidovým mostíkom.
Zvažované hormóny produkujú rad biologických účinkov: pre stimuláciu transportu vody a solí cez membránu majú presorické účinky, zvyšujú kontrakcie hladkého svalstva maternice pri pôrode, zvyšuje sekréciu mliečnych žliaz. Treba poznamenať, že vazopresín má antidiuretický účinok vyšší ako oxytocín, zatiaľ čo druhý ovplyvňuje maternicu a mliečnu žľazu. Hlavným regulátorom sekrécie vazopresínu je príjem vody, v obličkových kanáloch sa viaže na receptory v cytoplazmatických membránach s následnou aktiváciou enzýmu adenylátcyklázy v nich. Pre väzbu hormónu na receptor a pre biologický účinok sú zodpovedné rôzne časti molekuly.
Hypofýzy spojené s hypotalame cez nervový systém, kombinuje celé číslo funkčné endokrinného systému podieľať na zabezpečenie stálosti vnútorného prostredia (homeostázy). Vnútri endokrinné homeostatickej regulácia je založená na princípe spätnej väzby medzi predného laloku hypofýzy a zhelezami- "ciele" (štítnej žľazy, kôry nadobličiek, pohlavné žľazy). Prebytok hormón zhelezoy- "cieľ", pomaly, a jeho nedostatok stimuluje sekréciu a izoláciu zodpovedajúcu tropického hormónu. Systém spätnej väzby zahŕňa hypotalamus. To je v nej citlivé na hormóny železných cieľov, receptorových zón. Špecificky sa viazať na cirkulujúcich hormónov v krvi a menia reakcie v závislosti od koncentrácie hormónov hypotalamu receptory prenášajú svoj účinok na príslušné hypotalamických centier, ktoré koordinujú činnosť predného laloku hypofýzy, hypotalamu uvoľňujúce hormóny adenogipofizotropnye. Preto by hypotalamus mal byť považovaný za neuroendokrinný mozog.